

Cytosinstruktur, funksjoner, egenskaper, syntese
- 2800
- 49
- Theodor Anders Hopland
De Cytosin Det er en pyrimidin nitrogenbase, som tjener for biosyntesen av cytidin-5'-monofosfat og dexitidin-5'-monofosfat. Disse forbindelsene tjener til henholdsvis biosyntese fra deoksyribonukleinsyre (DNA) og ribonukleinsyre (RNA). DNA lagrer genetisk informasjon og RNA har forskjellige funksjoner.
Hos levende vesener finnes ikke cytosin fritt, men ofte ribonukleotider eller deoksyribonukleotider. Begge typer forbindelse har en fosfatgruppe, en ribose og en nitrogenbase.
Kilde: VESPROM [Public Domain] Karbon 2 av ribosen har en okshydrillo (-oh) gruppe i ribonukleotidene, og et hydrogenatom (-h) i deoksyribonukleotidene. Avhengig av antall fosfatgrupper som er til stede, er det Citidín-5'-monofosfat (CMP), Citidín-5'-difosfat (CDP) og Citidín-5'-triffosfat (CTP) (CTP).
Desoxygenated ekvivalenter kalles desoxicitidin-5'-monofosfat (DCMP), desoxicitidin-5'-difosfat (DCDP) og desoxicitidin-5'-triffosfat (DCTP).
Cytosin deltar, i sine forskjellige former, deltar i forskjellige funksjoner, for eksempel biosyntesen av DNA og RNA, biosyntesen av glykoproteiner og regulering av ekspresjonen av genet.
[TOC]
Struktur og egenskaper
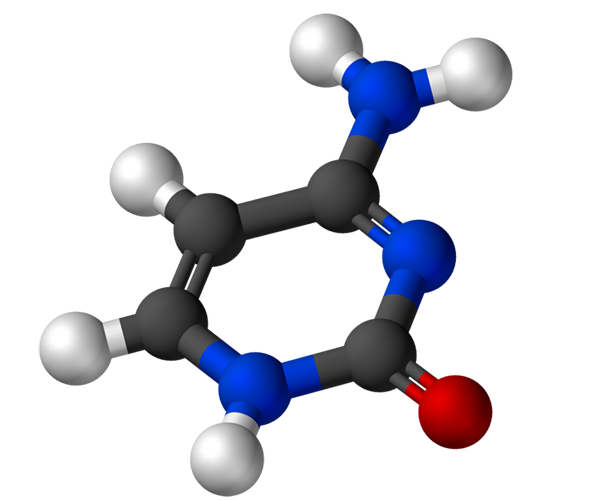
Cytosinet, 4-amino-2-hydroxipirimidin, har den empiriske formelen C4H5N3Eller, hvis molekylvekt er 111,10 g/mol, og renses som et hvitt pulver.
Strukturen til cytosinet er en heterocyklisk, aromatisk og plan ring. Maksimal absorbansbølgelengde (ʎMaks) Det er 260 nm. Fusjonstemperaturen til cytosinet overstiger 300 ºC.
For å danne et nukleotid binder cytosin kovalent, gjennom nitrogen 1, ved hjelp av måter. Carbon 5 'er esterifisert med en fosfatgruppe.
Biosyntese
Pyrimidin -nukleotidbiosyntese har en felles vei, som består av seks trinn katalysert av enzymer. Veien begynner med karbamoil-fosfatbiosyntese. I prokaryoter er det bare ett enzym: karbamoilfosfatsyntase. Dette er ansvarlig for syntese av pyrimidiner og glutamin. I eukaryoter er det karbamoilfosfatsyntase I og II, som er henholdsvis ansvarlige for glutamin og pyrimidinbiosyntese.
Det andre trinnet består av dannelsen av N-karbamoilaspartato, fra Carboil fosfat og aspartat, reaksjon katalysert av transpartatet Transcabamoilasa (ATCASA).
Det tredje trinnet er syntesen av L-dihydrotoate, som produserer lukking av pyrimidinringen. Dette trinnet blir katalysert av dihydrootase.
Det fjerde trinnet er dannelsen av Orotato, som er en redoksreaksjon katalysert av dihydroorotatdehydrogenase.
Det femte trinnet består av dannelsen av goldylat (OMP) ved bruk av fosforibosylpyrofosfat (PRPP) som et underlag, og fosforibosylorotatoverføringen som katalysator som katalysator.
Det sjette trinnet er dannelsen av uridilaato (uridin-5'-monofosfat, UMP), reaksjon katalysert av en OMP-decarboxylase.
Det kan tjene deg: Flora og fauna fra HidalgoFølgende trinn består av fosforylering av UMP, katalysert av kinaser, for å danne UTP, og overføring av en aminogruppe fra glutamin til UTP for å danne CTP, reaksjon katalysert ved CTP -syntetase.
Biosynteseforordning
Hos pattedyr finner regulering sted ved karbamoilfosfatsyntase.
Carbamoil Syntasa II er regulert av negativ tilbakemelding. Dets regulatorer, UTP og PRPP, er henholdsvis hemmer og aktivator av dette enzymet.
I vev som ikke er lever, er karbamoilfosfatsyntase II den eneste kilden til karbamoilfosfat. Mens i leveren, under forhold med overflødig ammoniakk, produserer karbamoilfosfatsyntase I, i mitokondriene, karbamoilfosfat, som transporteres til cytosol, hvor den kommer inn i biosyntesen av pyrimidin.
Et annet reguleringspunkt er OMP-Discarboxylase, som er regulert av konkurrerende hemming. Produktet av reaksjonen, UMP, konkurrerer med OMP for bindingsstedet i OMP-Descarboxilasa.
Pyrimidiner, som cytosin, blir resirkulert
Gjenvinning av pyrimidin har som en funksjon gjenbruk av pyrimidiner uten behov for Novo -biosyntese, og unngå nedbrytningssti. Gjenvinningsreaksjonen katalyseres av fosforibosyltransferase pyrimimidin. Den generelle reaksjonen er som følger:
Pyrimidin + PRPP -> Nukleosidpyrimidin 5 '-Monofosfat + PPI
I virveldyr finnes fosforibosyltransferase -pyrimimidin i erytrocytter. Pyrimidin -underlaget til dette enzymet er uracil, timina og orotato. Cytosin blir indirekte resirkulert fra uridin-5'-monofosfat.
Funksjon i DNA -biosyntese
Under DNA -replikasjon kopieres informasjonen i DNA i DNA gjennom en DNA -polymerase.
RNA -biosyntese trenger deoksynukleotider tryfosfat (DNTP), nemlig: Dexitimid Tryphosphate (DTTP), Dexicitidy Triffosphate (DCTP), de -alxyxyadenin Tryphyse (DATP) og Dexiguanin Tryphosphate (DDGTPYSE (DATP). Reaksjonen er:
(DNA)n avfall + Dntp -> (DNA)N+1 Avfall + PPI
Uorganisk pyrofosfat (PPI) hydrolyse gir energi for biosyntesen av RNA.
Funksjon i stabiliseringen av strukturen til DNA
I den doble DNUS -helixen er en purin, av en kjede, knyttet til pyrimidin, av den motsatte kjeden, ved hydrogenbindinger. Dermed er cytosin alltid knyttet til guanin av tre hydrogenbindinger: adenin er knyttet til timin av to hydrogenbindinger.
Det kan tjene deg: viktigheten av biologiHydrogenbindinger brytes når en renset naturlig DNA -løsning ved pH 7, blir utsatt for temperaturer større enn 80 ºC. Dette får den doble DNA -propellen til å danne to separate kjeder. Denne prosessen er kjent som denaturering.
Temperaturen som 50% av DNA er denaturert er kjent som smeltetemperaturen (TM). DNA -molekyler hvis guanin og cytosin.
Ovennevnte er den eksperimentelle testen at et større antall hydrogenbindinger bedre stabiliserer de naturlige DNA -molekylene.
Funksjon av rike regioner i cytosin i DNA
Nylig ble det funnet at DNA fra den menneskelige cellekjernen kan ta i bruk interkalerte motivstrukturer (IM). Disse strukturene er produsert i cytosinregioner.
IM -strukturen består av fire DNA -kjeder, i motsetning til det klassiske Double Propell DNA som har to kjeder. Mer spesifikt er to parallelle duplekskjeder ispedd en antiparallellorientering, og opprettholdes sammen av et par hemiprotonerte cytosiner (C: C: C+).
I det menneskelige genomet finnes IM -strukturer i regioner som promotorer og telomerer. Antallet IM -strukturer er høyere i løpet av G1/S -fasen av cellesyklusen, der transkripsjonen er høy. Disse regionene er proteinsjenkjenningssider som er involvert i aktivering av transkripsjonsmaskiner.
På den annen side, i regionene som er rike på guaninbaser (c) på rad, har DNA en tendens til å ta i bruk propellformen A, under dehydreringsforhold. Denne formen er typisk for RNA og dobbelt DNA-ARN blandede bånd under transkripsjon og replikasjon, og på bestemte tidspunkter når DNA er koblet til protein.
Det er vist at regionene med påfølgende cytosinbaser skaper en elektropositiv lapp i hovedspalte av DNA. Derfor antas det at disse regionene binder seg til proteiner, noe som disponerer visse genomiske regioner med genetisk skjørhet.
Funksjon i RNA -biosyntese
Under transkripsjonen kopieres informasjonen i DNA i RNA gjennom et Pon -polymerase -RNA. RNA -biosyntese trenger tryposfatnukleosider (NTP), nemlig: Tryphyse citidin (CTP), tryposfat uridin (UTP), adenin -tryposfat (ATP) og tryphosfat guanin (GTP). Reaksjonen er:
Kan tjene deg: Flora og Fauna of Campeche: Representative arter(RNA)n avfall + NTP -> (RNA)N+1 Avfall + PPI
Uorganisk pyrofosfat (PPI) hydrolyse gir energi for biosyntesen av RNA.
Funksjon i biosyntese av glykoproteiner
Den sekvensielle overføringen av heksoser for å danne oligosakkarider, o-lettater til proteiner, skjer fra nukleotidforløpere.
Hos virveldyr består det siste trinnet i biosyntesen av O-ligerte oligosakkarider i tilsetning av to sinaltyreavfall (N-acetylneuramin) fra en forløper av citidin-5'-monofosfat (CMP). Denne reaksjonen er produsert i transsekken til Golgi.
Cytosin- og cellegiftbehandlinger mot kreft
Tetrahydrofolatyre (FH4) er en gruppe -ch -kilde3, Og det er nødvendig for DTMP -biosyntese fra dump. FH2 er også dannet. FH2 -reduksjon til FH4 krever folat- og NADPH -reduktase. Noen folatreduktaseinhibitorer, for eksempel aminopterin og metretothrexate brukes i kreftbehandling.
Metotrexan er en konkurransedyktig hemmer. Folatreduktasen følger med 100 ganger mer affinitet til denne hemmeren enn til underlaget. Aminapterin fungerer på samme måte.
Inhiberingen av folatreduktase hindrer indirekte DTMP -biosyntese, og derfor den av DCTP. Direkte hemming skjer gjennom hemmere av timidilate syntetase, som katalyserer DTMP fra dump. Disse hemmere er 5-fluorouracil og 5-fluoro-2-dioxiuridin.
For eksempel er 5-fluoroacil ikke en hemmer, men blir først gjenvinningsveien, i deoxyurid mofosfat D (Fdump), som binder seg til timidilatesyntasasen og hemmer det.
Stoffer analogt med glutamin, azaserin og acivicin, hemmer amidotransferase glutamin. Azarine var et av de første oppdagede stoffene som fungerer som selvmord inaktiverende.
Referanser
- Assi, h.TIL., Garavís, m., González, ca., Og Damha, m.J. 2018. I-Motif DNA: Strukturelle trekk og mening til cellebiologi. Nuclei Acids Research, 46: 8038-8056.
- Bohinski, r. 1991. Biokjemi. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokjemi. Redaksjonell reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, p., Baltimore, d., Darnell, J. 2003. Cellulær og molekylær biologi. Redaksjonell Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Mexico, Sāo Paulo.
- Nelson, d. L., Cox, m. M. 2008. Lehninger-prinsipper for biokjemi. W.H. Freeman, New York.
- Voet, d. og Voet, J. 2004. Biokjemi. John Wiley og sønner, USA.