

Chloropasts egenskaper, funksjoner og struktur
- 1130
- 119
- Dr. Andreas Hopland
De Kloroplaster De er en type celleorganeller avgrenset av et komplekst membransystem, karakteristisk for planter og alger. I dette plastidiet er klorofyll, pigment som er ansvarlig for prosessene med fotosyntesen, grønnsaker, grønt av grønnsaker og tillater det autotrofiske livet til disse avstamningene.
I tillegg er kloroplaster relatert til generering av metabolsk energi (ATP - adenosin tryfosfat), syntese av aminosyrer, vitaminer, fettsyrer, lipidkomponenter i deres membraner og reduksjon av nitritter. Det har også en rolle i produksjonen av forsvarsstoffer mot patogener.
Kloroplast. Av Miguelsierra [GFDL (http: // www.gnu.Org/copyleft/fdl.HTML) eller CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons Denne fotosyntetiske organellen har sitt eget sirkulære genom (DNA), og det foreslås at de, i likhet med mitokondrier, stammer fra en symbioseprosess mellom en vert og en forfedre fotosyntetiske bakterier.
[TOC]
Opprinnelse
Kloroplaster er organeller som har kjennetegn på grupper av veldig fjerne organismer: alger, planter og prokaryoter. Dette beviset tyder på at organelle stammer fra et prokaryotisk organ med evnen til å utføre fotosyntese.
Det anslås at den første eukaryote organismen, med evnen til å utføre fotosyntese, oppsto omtrent 1.000 millioner år. Testene indikerer at dette viktige evolusjonære spranget var forårsaket av anskaffelse av en cyanobakteriell av en eukaryotisk gjest. Denne prosessen ga opphav til forskjellige linjer med røde, grønne og planter alger.
Tilsvarende reises sekundære og tertiære symbiosehendelser der en eukaryotisk avstamning etablerer et symbiotisk forhold til en annen fotosyntetisk eukaryota i fritt liv.
I løpet av evolusjonen har genomet til den antatte bakterien blitt redusert, og noen av dens gener er blitt overført og integrert i kjernenes genom.
Organiseringen av det nåværende kloroplastgenomet husker det av en prokaryotisk, men det har også attributter til eukaryotisk genetisk materiale.
Endosimbiotisk teori
Endosimbiotisk teori ble foreslått av Lynn Margulis i en serie bøker utgitt mellom 60- og 80 -tallet. Imidlertid var det en idé som allerede hadde kjørt siden 1900 -tallet, foreslått av Mereschkowsky.
Denne teorien forklarer opprinnelsen til kloroplaster, mitokondrier og basale kropper som er til stede i Scoures. I følge denne hypotesen var disse strukturene en gang gratis prokaryoter.
Det er ikke mye bevis som støtter endosimbiotisk opprinnelse til basale kropper fra mobile prokaryoter.
Derimot er det viktige bevis som støtter endosimbiotisk opprinnelse til mitokondrier fra α-proteobakterier og kloroplaster fra cyanobakterier. Det klareste og sterkere beviset er likheten mellom begge genomene.
Generelle kjennetegn ved kloroplaster
Kloroplaster er den mest iøynefallende typen plastider av planteceller. De er ovale strukturer omgitt av membraner, og inne i den oppstår den mest berømte prosessen med autotrofe eukaryoter: fotosyntese. De er dynamiske strukturer og har sitt eget genetiske materiale.
De er vanligvis lokalisert på planters blader. En typisk plantecelle kan ha 10 til 100 kloroplaster, selv om antallet er ganske variabelt.
I likhet med mitokondriene, oppstår arven etter kloroplaster av foreldre til barn av en av foreldrene og ikke av begge deler. Faktisk er disse organellene ganske like mitokondrier i forskjellige aspekter, selv om de er mer komplekse.
Struktur (deler)
Kloroplast. Av GMSOTAVIO [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0) eller GFDL (http: // www.gnu.Org/copyleft/fdl.html)], fra Wikimedia Commons Kloroplaster er store organeller, 5 til 10 um lengde. Egenskapene til denne strukturen kan visualiseres under et tradisjonelt optisk mikroskop.
Kan tjene deg: nukleosomDe er omgitt av en dobbel lipidmembran. I tillegg har de et tredje indre membransystem, kalt Tilacoid -membraner.
Dette siste membraniske systemet danner et sett med strukturer som ligner på et album, kjent som Tilacoides. Foreningen av thilacoider i batterier kalles "grana" og er koblet til hverandre.
Takket være dette trippelmembransystemet er kloroplastens indre struktur sammensatt og er delt inn i tre mellomrom: intermembranområdet (mellom de to ytre membranene), stroma (funnet i kloroplasten og utenfor membranen til tililacoid) og av Siste lumen til tilacoid.
Eksterne og indre membraner
Membransystemet er relatert til ATP -generasjon. Som mitokondriemembraner er det den indre membranen som bestemmer passering av molekyler inne i organellen. Fosfaditilkolin og fosfaditetslglycerol er de mest tallrike lipidene av kloroplastmembraner.
Den ytre membranen inneholder en serie porer. Små molekyler kan legge inn disse kanalene fritt. Den indre membranen tillater i mellomtiden ikke fri transitt av denne typen molekyler med lav vekt. For at molekylene skal komme inn, må de gjøre det ved hjelp av spesifikke transportører forankret til membranen.
I noen tilfeller er det en struktur som kalles perifert retikulum, dannet av et membrannettverk, spesielt stammer fra den indre kloroplastmembranen. Noen forfattere anser dem som unike for planter med C4 -metabolisme, selv om de er funnet i C3 -planter.
Funksjonen til disse tubuli og vesikler er ennå ikke klar. Det foreslås at de kan bidra til rask transport av metabolitter og proteiner i kloroplast eller for å øke overflaten til den indre membranen.
Tilacoidmembran
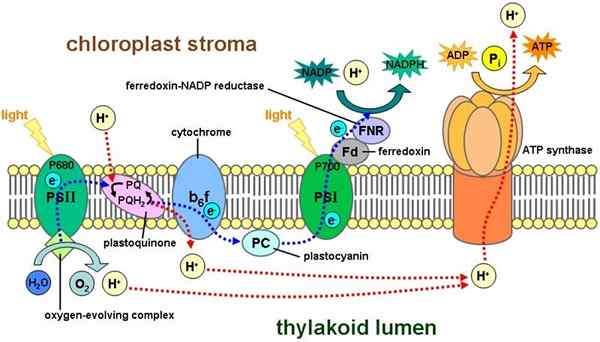
Tilacoidmembran. Tameria Sur Wikipédia Anglais [Public Domain], via Wikimedia Commons Elektron transportørkjeden involvert i fotosyntetiske prosesser skjer i dette membransystemet. Protonene pumpes gjennom denne membranen, fra stroma til det indre av tilacoides.
Denne gradienten resulterer i ATP -syntese, når protoner blir rettet igjen til stroma. Denne prosessen tilsvarer den som oppstår i den indre membranen til mitokondriene.
Tilacoidmembranen dannes av fire typer lipider: monogalaktosyldiacylglycerol, diglaktosyldiacylglyserol, sulfokinovosyldiacylglycerol og fosfatidylglyserol. Hver type oppfyller en spesiell funksjon i lipid -dobbeltlaget i denne delen.
Tilacoid
Tilacoider er membranøse strukturer i form av poser eller flate plater som er stablet i en "Cochineal”(Flertallet av denne strukturen er Granum). Disse albumene har en diameter på 300 til 600 nm. I det indre rommet til tilacoid kalles det lumen.
Tilacoid -stablingsarkitektur er fortsatt diskutert. Det foreslås to modeller: den første er den spiralformede modellen, der tilacoidene rulles mellom pogging i form av en propell.
I kontrast foreslår den andre modellen en bifurkasjon. Denne hypotesen antyder at grana er dannet av stroma bifurkasjoner.
Stroma
Stroma er den gelatinøse væsken som omgir tilacoid og finnes i det indre området i kloroplasten. Denne regionen tilsvarer cytosolen til de antatte bakteriene som oppsto denne typen plastidium.
I dette området er DNA -molekyler og en stor mengde protein og enzymer. Spesielt er enzymene som deltar i Calvin -syklusen, for fiksering av karbonanhydrid i den fotosyntetiske prosessen. Du kan også finne stivelsesgranuler
Kan tjene deg: cytoplasma: funksjoner, deler og egenskaperI stroma er ribosomene til kloroplaster, siden disse strukturene syntetiserer sine egne proteiner.
Genom
En av de viktigste egenskapene til kloroplaster er at de har sitt eget genetiske system.
Det genetiske materialet til kloroplaster består av sirkulære DNA -molekyler. Hver organelle har flere kopier av dette sirkulære molekylet på 12 til 16 kb (kilobaser). De er organisert i strukturer som kalles nukleoider og består av 10 til 20 kopier av plastgenomet, sammen med proteiner og RNA -molekyler.
Kloroplast DNA koder for omtrent 120 til 130 gener. Disse resulterer i proteiner og RNA relatert til fotosyntetiske prosesser som komponenten i fotosystem I og II, ATP -syntasen og en av Rubisco -underenhetene.
Rubisco (ribulosa-1,5-biskoposfat karboksylase/oksygenase) er et avgjørende enzymatisk kompleks i Calvin-syklusen. Faktisk vurderes det mest tallrike proteinet på planeten Jorden.
Overfør RNA og ribosomales brukes i oversettelsen av meldinger som er kodet i kloroplastgenomet. Inkluderer ribosomales 23s, 16s, 5s og 4,5s og 30 overføringsribber. Det koder også for 20 ribosomale proteiner og visse underenheter av RNA -polymerase.
Imidlertid er visse elementer som er nødvendige for driften av kloroplasten kodet i kjernefysisk kjernefysisk genom.
Funksjoner
Kloroplaster kan betraktes som viktig metabolsk senter i planter, der flere biokjemiske reaksjoner oppstår takket være det brede spekteret av enzymer og proteiner forankret til membraner som disse organellene inneholder.
De har en kritisk funksjon i planteorganismer: det er stedet der fotosyntetiske prosesser oppstår, der sollys blir omdannet til karbohydrater, med oksygen som sekundært produkt.
I kloroplaster også en serie sekundære funksjoner av biosyntese. Neste vil vi diskutere hver funksjon i detalj:
Fotosyntese
 Fotosyntese (til venstre) og pust (DCHA). Bilde av høyre hentet fra BBC
Fotosyntese (til venstre) og pust (DCHA). Bilde av høyre hentet fra BBC Fotosyntesen oppstår takket være klorofyll. Dette pigmentet er innenfor kloroplastene, i tilacoidmembranene.
Den er sammensatt av to deler: en ring og en hale. Ringen inneholder magnesium og er ansvarlig for absorpsjonen av lys. Den kan absorbere blått og rødt lys, og gjenspeiler den grønne sonen i lysspekteret.
Fotosyntetiske reaksjoner oppstår takket være elektronoverføring. Energi fra lys gir energi til klorofyllpigment (det sies at molekylet er "begeistret av lys"), noe som forårsaker en bevegelse av disse partiklene i tilacoidmembranen. Klorofyll oppnår elektronene fra et vannmolekyl.
Denne prosessen resulterer i dannelse av en elektrokjemisk gradient som tillater syntese av ATP i stroma. Denne fasen er også kjent som "lysende".
Den andre delen av fotosyntesen (eller mørk fase) forekommer i stroma og fortsetter i cytosolen. Det er også kjent som karbonfikseringsreaksjoner. På dette stadiet brukes produktene fra ovennevnte reaksjoner til å bygge karbohydrater fra CO2.
Biomolekyler syntese
I tillegg har kloroplaster andre spesialiserte funksjoner som tillater utvikling og vekst av anlegget.
I denne organellen oppstår assimilering av nitrater og sulfater, og de har de nødvendige enzymer for syntese av aminosyrer, fytohormoner, vitaminer, fettsyrer, klorofyll og karotenoider.
Kan tjene deg: Integriner: Kjennetegn, struktur og funksjonerVisse studier har identifisert et viktig antall aminosyrer syntetisert av denne organellen. Kirk og samarbeidspartnere studerte produksjonen av aminosyrer i kloroplastene til Vicia faba L.
Disse forfatterne fant at de mest tallrike syntetiserte aminosyrene var glutamat, aspartat og treonin. Andre typer, for eksempel Alanina, Serine og Glycina ble også syntetisert, men i mindre mengde. De resterende aminosyrene ble også påvist.
Ulike gener involvert i lipidsyntese er blitt isolert. Kloroplaster har de nødvendige veiene for syntese av isaprenoid lipider, viktige for klorofyllproduksjon og andre pigmenter.
Patogenforsvar
Planter har ikke et immunforsvar utviklet som dyr for dyr. Derfor må cellulære strukturer produsere antimikrobielle stoffer for å kunne forsvare seg mot skadelige midler. For dette formålet kan planter syntetisere reaktiv oksygen (ROS) eller salisylsyrearter.
Kloroplaster er relatert til produksjonen av disse stoffene som eliminerer mulige patogener som kommer inn i planten.
De fungerer også som en "molekylære sensorer" og deltar i varslingsmekanismer, og formidler informasjon til andre organeller.
Andre plastider
Kloroplaster tilhører en familie av vegetabilske organeller kalt plastider eller plast. Kloroplaster skiller seg hovedsakelig fra resten av plastidene ved å ha klorofyllpigment. De andre plastidene er:
-Chromoplastos: Disse strukturene inneholder karotenoider, er til stede i blomster og blomster. Takket være disse pigmentene har plantestrukturer gule, oransje og røde farger.
-Leukoplastene: Disse plastidene inneholder ikke pigmenter og er derfor hvite. De fungerer som en reservasjon og finnes i organer som ikke får direkte lys.
-Amyloplastene: De inneholder stivelse og finnes i røtter og knoller.
Plastidene stammer fra strukturer som kalles protoplast. En av de mest overraskende egenskapene til plastider er deres eiendom til å endre type, selv om de allerede er i modent stadium. Denne endringen utløses av miljømessige eller iboende signaler av planten.
For eksempel er kloroplaster i stand til å gi opphav til kromoplaster. For denne endringen blir tilacoidmembranen desintegrates og karotenoider syntetiseres.
Referanser
- Allen, J. F. (2003). Hvorfor kloroplaster og mitechondria inneholder genomer. Sammenlignende og funksjonell genomikk, 4(1), 31-36.
- Cooper, g. M (2000). Cellen: tilnærming molekylær. Andre utgave. Sinaauer Associates
- Daniell, h., Lin, c.-S., Yu, m., & Chang, w.-J. (2016). Kloroplastgenomer: Mangfold, evolusjon og anvendelser innen genteknologi. Genombiologi, 17, 134.
- Gracen, v. OG., Hilliard, J. H., Brown, r. H., & West, S. H. (1972). Periphal Resticulum i kloroplaster av planter som er forskjellige i CO 2 -fiksering av patoways og hutortespirion. Anlegg, 107(3), 189-204.
- Gray, m. W. (2017). Lynn Margulis og endosymbionnt -hypotesen: 50 år senere. Biologi av cellemolekylæren, 28(10), 1285-1287.
- Jensen, p. OG., & Leister, D. (2014). Kloroplast evolusjon, struktur og funksjoner. F1000 Prime -rapporter, 6, 40.
- Kirk, p. R., & Leech, r. M. (1972). Aminosyrebiosyntese av isolerte kloroplats under fotosyntese . Plantefysiologi, femti(2), 228-234.
- Kobayashi, k., & Wada, h. (2016). Lipiders rolle i kloroplastbiogenese. I Lipider i plante- og algeutvikling (s. 103-125). Springer, Cham.
- Sowden, r. G., Watson, s. J., & Jarvis, P. (2017). Kloroplastens rolle i plantepatologi. Essays in Biochemistry, EBC20170020.
- Klokere. R., & Hoober, j. K. (2007). Strukturen og funksjonen til plastider. Springer Science & Business Media.
- « Demoseklassifisering, egenskaper, habitat, eksempler
- Hexactinélidos klassifisering, egenskaper, habitat, arter »