

Ioniske kanaler Struktur, funksjoner, typer
- 3967
- 1029
- Magnus Sander Berntsen
De ioniske kanaler De er hule membranale strukturer som danner kanaler eller porer som krysser membranens tykkelse og kommuniserer utsiden av cellen med dens cytosol og omvendt; Noen kan ha et portsystem som regulerer åpningen.
Disse kanalene er fulle av vann og kontrollerer passering av spesifikke ioner fra den ene siden til den andre av membranen. De er dannet av proteiner som er typiske for cellemembraner som danner sylindriske rør -formede strukturer som går gjennom dem.
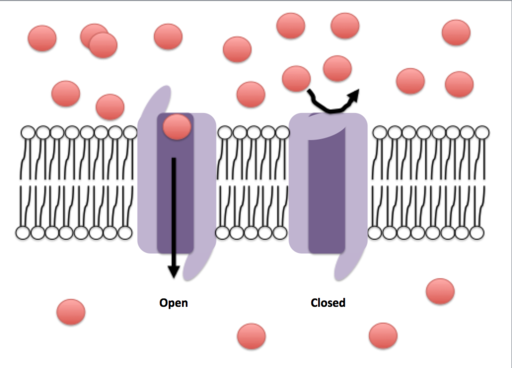
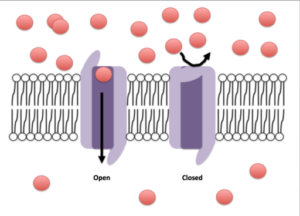 Åpen og lukket konformasjon av en ionisk kanal (kilde: Efazzari [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Åpen og lukket konformasjon av en ionisk kanal (kilde: Efazzari [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Transportmekanismer gjennom disse membranene kan klassifiseres mye i passiv eller aktiv transport. Forpliktelsene er de som tillater passering av stoffer til fordel for konsentrasjonsgradientene, i mellomtiden krever eiendelene energiforbruk, siden de fortrenger stoffer mot konsentrasjonsgradientene.
Ioniske kanaler utgjør en passiv transportmekanisme som kan klassifiseres i henhold til deres spesifisitet, det vil si i henhold til den type ion de lar passere, eller avhengig av måten de åpner eller lukker eller lukker.
https: // giphy.com/gifs/ion-channel-y07c7ocoigykjl6tzz
Hovedfunksjonen til disse membranale transportsystemene er å tillate den regulerte passering av stoffer i eller utenfor cellene og dermed opprettholde intracellulære konsentrasjoner av ioner og andre stoffer.
Tilstedeværelsen av cellemembraner og ioniske kanaler fundamentalt for å opprettholde konsentrasjonsforskjeller mellom intracellulære og ekstracellulære medier, som har relevans fra mange synspunkter.
Ioniske kanaler, spesielt de avhengige ligandene, er veldig viktige innen farmakologi og medisin, siden mange medisiner kan etterligne funksjonene til naturlige ligander og bli med i kanalen, åpne eller lukke den, ettersom tilfellet kan være tilfelle.
Andre medisiner er i stand til å blokkere unionsstedet og dermed forhindre virkningen av naturlig ligand.
[TOC]
Struktur
Strukturen til ioniske kanaler dannes av spesifikke transmembranproteiner som har en rørformet form og etterlater en pore eller hull som tillater kommunikasjon mellom interiøret og utsiden av cellen eller mellom intracellulære rom (organeller).
Hver ioniske kanal innebærer et spesifikt strukturelt protein av membranen og mer enn 100 gener som koder for spesifikke ioniske kanaler er beskrevet.
For natriumkanalen, for eksempel, 10 gener som kalles SCN som koder for forskjellige proteiner fordelt i forskjellige vev med spesielle funksjoner og strukturer.
På samme måte en betydelig mengde gener som kodifiserer forskjellige proteiner som utgjør kaliumkanaler som tilhører forskjellige familier og har forskjellige mekanismer for aktivering, åpning og inaktivering.
Proteinstruktur av en ionisk kanal
Vanligvis er en funksjonell ionisk kanal assosiert med en membran sammensatt av monteringen av 4 til 6 lignende polypeptidunderenheter (HOMO oligomerer) eller forskjellige (hetero oligomerer) som danner en sentral pore mellom dem.
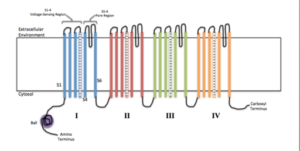 Skjema for membranale underenheter av en ionisk kanal (kilde: Efazzari [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Skjema for membranale underenheter av en ionisk kanal (kilde: Efazzari [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Hver underenhet varierer i henhold til kanalens egenskaper og egenskaper, siden mange er spesifikke for visse ioner og har forskjellige åpnings- og lukkemekanismer.
Kan tjene deg: cnidocytter: egenskaper, struktur, funksjoner, typerNoen kanaler består av en enkelt polypeptidkjede organisert i gjentatte motiver som går gjennom membrantykkelsen og funksjonen som ekvivalent til et proteinunderenhet flere ganger.
I tillegg til disse underenhetene, som er kjent i litteratur som α -underenheter, har noen ioniske kanaler også en eller flere hjelpeunderenheter (ß eller γ) som regulerer åpningen og lukkingen av dem.
Spesifisiteten til hver kanal er relatert til diameteren til porene dannet av de transmembranale proteiner og sidekjedene (─R) til aminosyrene som utgjør dem.
På denne måten er det kanaler som bare savner natrium, kalium, kalsiumioner og så videre, fordi sidekjedene fungerer som "sil".
Ytterligere strukturelle egenskaper
Et annet viktig trekk ved mange kanaler er portene. Kanalene med disse egenskapene kan åpnes eller lukkes foran lokale endringer som oppstår i membranal mikroenking som omgir kanalen.
Avhengig av kanaltypen, kan disse endringene være mekaniske, termiske (temperaturendringer), elektrisk (spenningsendringer) eller kjemikalier (binding av en ligand).
I de såkalte passive ioniske kanalene, som er de som forblir åpne og tillater det spesifikke trinnet med bestemte ioner, har disse strukturene ikke porter eller er følsomme for ligander eller andre typer stimuli.
I andre ioniske kanaler, som er følsomme for nærvær eller tilkobling av ligander, er det et bindingssted for ligand enten på den ekstracellulære siden eller for cellcytosol, og i disse tilfellene har porene eller kanalene en port som kan åpnes eller lukkes I henhold til statusen til hans ligand.
Mekanisme for sekunder Messenger for åpning eller lukking av kanaler
Når det gjelder å ha et sted for ligand i den intracellulære delen, har disse kanalene vanligvis andre budbringere som ligander. Et eksempel på ioniske kanaler som åpner eller lukkes ved mekanismer for andre budbringere, er den av luktreseptorer:
Odoriferous molekyler binder seg til reseptorene sine på den ekstracellulære siden. Disse reseptorene er på sin side festet til et G -protein som aktiveres som på sin side aktiverer adenylciclase -proteinet som danner AMPC, som er en andre messenger.
AMPC blir med i et intracellulært veikrysssted for kalsiumkanaler, noe som resulterer i åpnings- og kalsiuminngang i cellen.
Som om det var en dominoeffekt, blir kalsium sammen med et sted for å koble en annen klorkanal, som genererer åpningen og avkjørselen til dette ionet, noe som forårsaker depolarisering av luktcellen.
Det er viktig å merke seg at endringene som genereres av ligandene eller stimuli som påvirker de ioniske kanalene, tilsvarer de konformasjonsendringene til proteinene som utgjør strukturen til kanalen.
Med andre ord er de konformasjonsendringene som kan flytte en port og lukke eller åpne en kanal ikke noe mer enn tilnærmingen eller distansen av proteinunderenhetene som utgjør den.
Andre aktiverings- og inaktiveringsmekanismer
Noen kanaler, spesielt spenningsavhengige kanaler, kan legge inn en ildfast tilstand der den samme spenningen endres som aktiverte dem nå ikke lenger aktivt dem.
Det kan tjene deg: Cellebiologi: Historie, hvilke studier, applikasjoner og konsepterFor eksempel, i spenningsavhengige kalsiumkanaler, åpner spenningsendringen kanalen og kalsium kommer inn, og når den er inne i cellen, blir det samme ionet sammen med et felles kanalkrysssted og den lukkes.
En annen form for reversibel inaktivering av kalsiumkanalen som forklarer dens ildfaste etter aktivering er parasforylering av kanalen på grunn av økningen i intern kalsiumkonsentrasjon.
Det vil si at en kalsiumkanal kan være irreversibelt inaktivert på grunn av tilstedeværelsen av patologisk høye konsentrasjoner av ionet, som medierer rekrutteringen av utfoldelsesenzymer av andre proteiner aktivert av kalsium.
Kanalene regulert av ligando kan inngå en ildfast tilstand når de blir forlenget til deres ligand, og mottar denne mekanismen.
Legemidler, giftstoffer og giftstoffer kan påvirke reguleringen av ioniske kanaler, lukke dem eller holde dem åpne eller i noen tilfeller okkupere ligandstedet og dermed forstyrre deres funksjon.
Funksjoner
Ioniske kanaler har mangfold av funksjoner, direkte eller indirekte.
- De er ansvarlige for å regulere strømmen av ioner gjennom plasma- og organelære membraner i alle celler.
- Tillat eksistensen av kontroll over de intracellulære konsentrasjonene av de forskjellige ionene.
- I nevroner og i muskelceller kontrollerer ioniske kanaler variasjonene i membranpotensialet som oppstår under handlingspotensialer og under post -synaptisk post -synaptiske potensialer til effektorceller.
- Kalsiumkanaler som genererer netto kalsiumstrømmer til intracellulært rom er ansvarlige for aktivering av mange enzymer og proteiner som deltar i mange metabolske prosesser.
- På samme måte initierer økningen i kalsium på grunn av en økning i transport mekanismen for frigjøring av nevrotransmittere til det synaptiske rommet til nevroner.
- Derfor er funksjonen til ioniske kanaler også relatert til cellulære kommunikasjonsmekanismer.
Generelt for transport gjennom membranen
Som nevnt ovenfor, kan de membranale transportmekanismene være aktive eller forpliktelser i henhold til eller ikke energien til cellen der de blir funnet. Passive mekanismer er klassifisert som enkel diffusjon og tilrettelagt diffusjon.
Enkel diffusjon
Enkel diffusjon tillater passasje gjennom fosfolipidstrukturen til småstørrede fett -oppløselige molekyler, med apolar og belastningsfrie funksjoner.
Således passerer for eksempel gasser som oksygen (O2) og karbondioksid (CO2), etanol og urea, for å nevne noen, til fordel for konsentrasjonsgradienten.
Tilrettelagt diffusjon
Den spredte diffusjonen er en som letter av protein, og denne passive transportmekanismen er det to typer: ionekanaler og transportproteiner eller transportproteiner.
Ioniske kanaler er den mest brukte mekanismen av ionetransportceller som ikke kan passere med enkel diffusjon, verken fordi de har elektrisk ladning og membranfosfolipider frastøter dem, på grunn av deres størrelse og polaritet eller andre karakteristiske.
Diffusjonen levert av transportproteiner brukes til transport av større stoffer med eller uten belastning, for eksempel glukose og annet sukker.
Kan tjene deg: grensesnittAktiv membranaltransport er en som oppstår mot konsentrasjonsgradienten til oppløsningen som transporteres og krever ATP -formet energiforbruk. Blant transportørene av denne typen er pumper og vesikulær transport.
Som et eksempel på pumpene er natrium/kalium, som tar tre brus og introduserer to kalium. Det er også kalsiumpumper.
https: // giphy.com/gifs/qsib5zeioyrufkuusb
Som et eksempel på vesikulær transport er endocytose, eksocytose, pinocytose og fagocytose; Alle disse aktive transportmekanismene.
Typer ionekanaler
Fra dette tidspunktet vil det bli referert til ioniske kanaler som tillater passering av ioner gjennom en membran til fordel for konsentrasjonsgradienter, det vil si at de er passive transportkanaler.
Generelt er hver av disse kanalene spesifikke for et enkelt ion, med forbehold om noen få kanaler som tillater transport av ionepar.
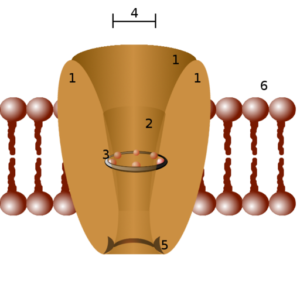 Strukturskjema for en ionisk kanal (kilde: outsslider (Paweł Tokarz) på PL.Wikipedia [Public Domain] via Wikimedia Commons)
Strukturskjema for en ionisk kanal (kilde: outsslider (Paweł Tokarz) på PL.Wikipedia [Public Domain] via Wikimedia Commons) En måte å klassifisere ioniske kanaler er å gruppere dem i henhold til mekanismen som er ansvarlig for åpningen. Således er passive kanaler, kanaler regulert av spenning (spenningsavhengige), kanaler regulert av ligando og kanaler regulert av mekaniske stimuli blitt beskrevet.
- Passive kanaler: De er kanaler som er permanent åpne og ikke svarer på noen form for stimulans; Disse er spesifikke for visse ioner.
- Spenningsavhengige kanaler: Disse kan åpnes eller lukkes (avhengig av kanalen) i møte med endringer i membranspenningen. De er veldig viktige for cellesignalering, spesielt i sentralnervesystemet til pattedyr.
- Lysavhengige kanaler: også kalt kanaler med koblingsdør eller regulert ved kobling, er vidt distribuert i de forskjellige kroppscellene til mennesker, men i nervesystemet utgjør de de ioniske kanalene aktivert av nevrotransmittere og er essensielle for synaptisk overføring og intercellulær skilting.
Eksempel på ligandavhengige ionekanaler aktivert av nevrotransmittere er natrium/kaliumkanaler aktivert av glutamat.
Aktivering av kolinergiske reseptorer, i dette tilfellet forening av acetylkolin til den postsynaptiske membranen (kanalligand), åpner ligandavhengige natriumkanaler og tillater inntreden av dette ionet etter konsentrasjonsgradienten.
- Kanaler regulert av mekaniske stimuli: De er kanaler som kan aktiveres ved distensjon eller trykk. Disse mekaniske kreftene overføres til kanalen gjennom cytoskjelettet og kanalen åpnes.
Referanser
- Bear, m. F., Connors, f. W., & Paradiso, m. TIL. (Eds.). (2007). Nevrovitenskap (Vol. 2). Lippinott Williams & Wilkins.
- Institutt for biokjemi og molekylær biofysikk Thomas Jesell, Siegelbaum, S., & HUDSPETH, A. J. (2000). Prinsipper for nevralvitenskap (Vol. 4, s. 1227-1246). OG. R. Kandel, J. H. Schwartz, & t. M. Jesell (red.). New York: McGraw-Hill.
- Lewis, ca. TIL., & Stevens, C. F. (1983). Acetylcholine Channel Ion -selektivitet: INS opplever et vandig miljø. Proceedings of the National Academy of Sciences, 80(19), 6110-6113.
- Nelson, d. L., Lehninger, a. L., & Cox, m. M. (2008). Lehninger prinsipper for biokjemi. Macmillan.
- Rawn, J. D. (1998). Biokjemi. Burlington, Massachusetts: Neil Patterson Publisher.
- Viana, f., av Peña, og., & Belmonte, C. (2002). Spesifisiteten til kald termotransduksjon bestemmes ved differensiell ionisk kanaluttrykk. Natur nevrovitenskap, 5(3), 254.
- « Flora kokoide egenskaper, sjangre, patologier, diagnose
- Trachycarpus -egenskaper, habitat, distribusjon, arter »