

Glioxylatsyklusegenskaper, reaksjoner, regulering, funksjoner
- 4140
- 536
- Thomas Karlsen
Han Glioxylatsyklus Det er en metabolsk rute som er til stede i planter, i noen mikroorganismer og i virvelløse dyr (fraværende i alle virveldyr), gjennom hvilke disse organismer kan omdanne fett til karbohydrater (sukker).
Denne ruten ble oppdaget i 1957, mens Kornberg, Krebs og Beevers prøvde å belyse hvordan bakterier som som som Escherichia coli De kunne vokse i nærvær av acetat som den eneste kilden til karbon, og hvordan frøplantene i spiring av Tártago (Ricinus communis) De kunne gjøre fettstoffene til karbohydrater.
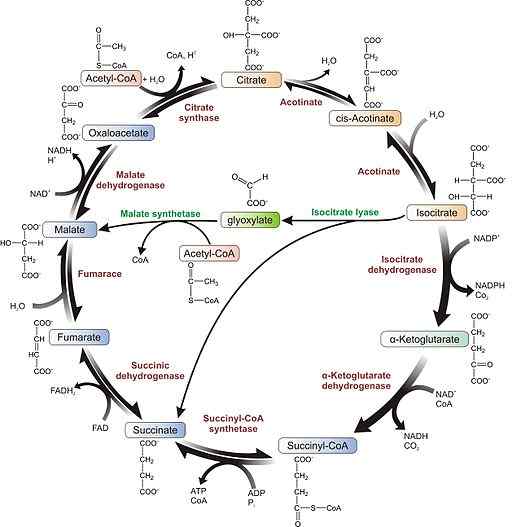
 Glioxilate Cycle Scheme (Kilde: Agrotman [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons)
Glioxilate Cycle Scheme (Kilde: Agrotman [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) Studiene av disse tre forskerne førte til oppdagelsen av to enzymer kjent som Liasa-isokitrat og den onde syntasen, som sammen med enzymene i Krebs-syklusen tillater succinatsyntese fra to acetyl-CoA-molekyler.
Den således produserte succinatet omdannes til malato gjennom trikarboksylsyresyklusen, og kan senere brukes til glukoseproduksjon ved glukoneogenese.
Denne ruten forekommer, i planter, i spesielle organeller som kalles glioxisomes og er avgjørende for overlevelse av frøplanter i de tidlige stadiene av spiring.
[TOC]
Kjennetegn
Glioxylatruten kan betraktes som en "modifisering" av Krebs -syklusen, med forskjellen at i den første ikke forekommer en oksidativ dekarboksylering, men at dikarboksylsyrer av fire karbonatomer kan dannes fra to -atomer fra to acetatenheter av to karbonatomer.
Denne egenskapen til glioxylatsyklusen er blitt beskrevet som en form som noen organismer må unngå (bypass ") tapet av karbonatomer i form av karbondioksid som identifiserer Krebs -syklusen.
Hos planter forekommer glioxylatsyklusen inne i noen cytosoliske organeller omgitt av en enkel membran som er kjent som glyxisomer. I andre organismer som gjær og alger, forekommer denne ruten i cytosolen i cytosolen.
Glioxisomer er strukturelt lik peroksisomer (noen forfattere anser dem som "spesialiserte peroksisomer"), andre organeller med ansvar for ß-oksidasjon av fettsyrer og eliminering av reaktive oksygenarter i eukaryote organismer.
Det kan tjene deg: gangliasidosInnvendig oksyderes fettsyrer for å produsere acetyl-CoA, som deretter kondenseres i forbindelser av fire karbonatomer. Disse forbindelsene blir selektivt transportert til mitokondriene, hvor de blir omdannet til Malato eller transporteres til cytosolen for å komme inn i den glukoneogene ruten (syntese av glukose).
Enzymer som deles mellom glioxylatruten og trikarboksylsyresyklusen eksisterer i mitokondrier og glioxisom som isoenzymer, noe som betyr at begge rutene fungerer mer eller mindre uavhengig uavhengig av den andre.
Glioxisomes forekomst
Glioxisomas er ikke til stede i plantevev permanent. De er spesielt rikelig under spiring av oleaginøse frø, som har liten fotosyntetisk kapasitet til å produsere karbohydratene de trenger for å vokse.
I fullt utviklede planter er deres deltakelse i fettmetabolisme ikke så viktig, siden sukker hovedsakelig oppnås ved fotosyntese.
Reaksjoner
Acetat fra nedbrytningen av fettsyrer fungerer som et rikt drivstoff i energi og som en kilde til fosfoenolpyruvat for glukosesyntese gjennom glukoneogenese. Prosessen skjer som følger:
Glioxilat syklus trinn
1- Glioksylatruten, lik den i Krebs-syklusen, begynner med kondensasjon av et acetyl-CoA-molekyl med et annet oksalacetat for å utføre citrat, reaksjon katalysert av enzymsitratsyntasen.
2- Aconitosa-enzym konverterer dette sitratet til isokitrate.
3- Isocitrato brukes som et underlag av liasa-isokitratenzymet for å danne succinatforbindelser og glioxylat.
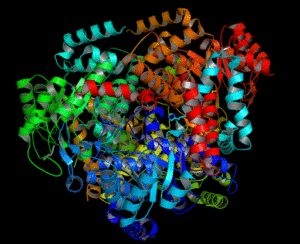 Molekylær struktur av Liasa -isokitratenzymet (kilde: Vrabiochemhw [CC0] via Wikimedia Commons)
Molekylær struktur av Liasa -isokitratenzymet (kilde: Vrabiochemhw [CC0] via Wikimedia Commons) 4- Glioxylatet tas av Malato Syntasa-enzymet for å produsere ondskap gjennom kondensasjon med et andre acetyl-CoA-molekyl.
Kan tjene deg: Organiske biomolekyler: Karakteristikker, funksjoner og eksempler5- Den onde blir omdannet til oksalacetat av den onde dehydrogenasen og sa at forbindelsen kan tjene som en forløper for den glukoneogene ruten eller kondensere med en annen acetyl-CoA for å starte syklusen på nytt igjen.
6- Succinatet som produseres kan også omdannes til fumarat og dette til Malato, og gir mer oksalacetatmolekyler for glukosedannelse. Ellers kan dette molekylet også eksporteres til mitokondriene for å jobbe i Krebs -syklusen.
Oksalacetat kommer inn i den glukoneogene ruten for glukoseproduksjon takket være den.
Regulering
Siden syklusene med glioxylat og trikarboksylsyrer deler mange mellommenn med hverandre, er det en koordinert forskrift mellom de to.
I tillegg er det nødvendig at det er kontrollmekanismer, siden syntesen av glukose og andre heksoser fra acetyl-CoA (fra fettnedbrytning) innebærer deltakelse på minst fire ruter:
- Β-oksidasjon av fettsyrer som produserer acetyl-CoA-molekyler som er nødvendige for både Krebs-syklusen, og at det i planter finner sted i glioxisomes.
- Glioxylatsyklusen, som også forekommer i glioxisomes og, som nevnt, produserer mellommenn som suksinat, ondskap og oksalacetat.
- Krebs -syklusen, som foregår i mitokondriene og der mellommennene saccinate, onde og oksalacetat også forekommer.
- Glukoneogenese, som oppstår i cytosol og tenker på bruk av oksalacetat omgjort til fosfoenolpyruvat for å syntetisere glukose.
Hovedkontrollpunktet er i enzymetisokitratdehydrogenase, hvis regulering innebærer en kovalent modifisering ved tilsetning eller fjerning av en fosfatgruppe.
Når enzymet er fosforylert, er det inaktivt, så isocytratet er rettet mot glukoseproduksjonsveien.
Funksjoner
For planter er glioxylatsyklusen grunnleggende, spesielt under spiringsprosessen, siden nedbrytningen av fett som er lagret i frøene, utnyttes for glukosesyntese i dårlig utviklet fotosyntetisk sett vev.
Kan tjene deg: glykogen: struktur, syntese, nedbrytning, funksjonerGlukose brukes som en kilde til å oppnå energi i form av ATP eller for dannelse av mer komplekse karbohydrater med strukturelle funksjoner, men noen av mellommennene som genereres under glioxylatruten kan også tjene synteseformålene til andre cellulære komponenter.
I mikroorganismer
Hovedfunksjonen til glioxylatsyklusen i mikroorganismer er å gi en "alternativ" metabolsk rute, slik at mikroorganismer er i stand til å dra nytte av andre karbon- og energikilder for deres vekst.
Slik er tilfellet med bakterier Escherichia coli, Når nivåene av noen formidlere av glykolyse og cytriskyresyklusen reduseres (isokitrat, 3-fosfoglyserat, pyruvat, fosfoenolpyruvat og oksalacetat) er enzymet) inhiBet og inhiberet i ishydrogenasen (som deltar i Krebs-syklyset) i ishydrogenasen (som deltar i Krebs) -hemmet og inhiBet og inhiBet og inhiBet, er inhiBet og inhiBet den som er inhiBet og inhiBet og inhiBet og inhiBet og inhiberte, er inhiBet og inhiBet og inhiBet og inhiBet den som er inhiBited og inhiBet og inhiBet, og er inhiBet og inhiBet. rettet mot glioksylatets rute.
Hvis denne ruten er aktiv på det tidspunktet bakteriene vokser i et medium rik på acetat, for eksempel, kan denne metabolitten brukes til å syntetisere karboksylsyrer av fire karbonatomer som senere kan stamme i dannelsen av energikarbohydrater.
For andre organismer som sopp, for eksempel, er det vist at patogenisitet i stor grad avhenger av tilstedeværelsen av en aktiv glioxylatsyklus, tilsynelatende av metabolske årsaker.
Referanser
- Dey, p., & Harborne, J. (1977). Plantebiokjemi. San Diego, California: Academic Press.
- Ensign, s. TIL. (2006). Gjennomgang av glyoksylatsyklusen: Alternative traséer for mikrobiell acetatassimilering. Mikrobiologi Molecular, 61 (2), 274-276.
- Garrett, r., & Grisham, C. (2010). Biokjemi (4. utg.). Boston, USA: Brooks/Cole. Cengage Learning.
- Lorenz, m. C., & Fink, g. R. (2001). Glyxylatsyklusen er nødvendig for soppvirulens. Natur, 412, 83-86.
- Mathews, c., Van Holde, K., & Ahern, K. (2000). Biokjemi (3. utg.). San Francisco, California: Pearson.
- Rawn, J. D. (1998). Biokjemi. Burlington, Massachusetts: Neil Patterson Publisher.
- Vallarino, J. G., & Osorio, S. (2019). Organiske syrer. I Posthavest Physiology and Biochemistry of Fruits and Vegetables (PP. 207-224). Elsevier Inc.
- « Mesohyppus -egenskaper, arter, reproduksjon, ernæring
- Paquimé kultur opprinnelse, beliggenhet, egenskaper, religion »