

Kromatintyper, egenskaper, struktur, funksjoner
- 1879
- 152
- Dr. Andreas Hopland
De Kromatin Det er komplekset som er dannet av DNA og proteiner, unike i eukaryote organismer. Når det gjelder proporsjoner inneholder det nesten dobbelt protein som av genetisk materiale. De viktigste proteiner i dette komplekset er histoner - små proteiner med positiv belastning som binder seg til DNA ved elektrostatiske interaksjoner. I tillegg har kromatin mer enn tusen proteiner forskjellige fra histoner.
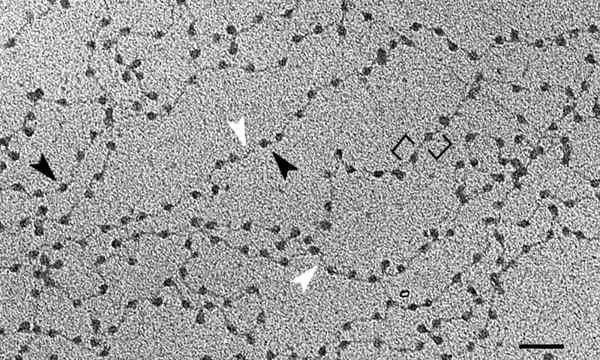
Den grunnleggende enheten av kromatin er nukleosom, som består av foreningen av histoner og DNA. Denne ordinasjonen minner beretningene om et halskjede. Etter å ha gått gjennom alle høyere organisasjonsnivåer, vil vi nå kromosomene.
Kilde: kromatin_nukleofilamenter.PNG: Chris Woodcockderivevativt arbeid: Gouttegd [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] Strukturen til kromatin er nært beslektet med kontrollen av genuttrykk. Det er to hovedtyper: eukromatin og heterokromatin.
Eukromatin er preget av å ha en lav grad av komprimering, noe som oversettes til høye transkripsjonsnivåer. I kontrast er heterokromatin transkripsjonelt inaktivt, på grunn av graden av komprimering så høy at det presenterer.
Strukturelt er det visse epigenetiske merker av histonene til begge typer kromatiner. Mens eukromatin er assosiert med acetylering, er heterokromatin assosiert med reduksjon i kjemiske grupper.
Det er noen heterokromatinregioner med strukturelle funksjoner, for eksempel telomerer og sentromerer.
[TOC]
Historisk perspektiv
Studien av genetisk materiale og dens strukturelle organisasjon begynner i 1831 når forsker Robert Brown beskriver kjernen. Et av de umiddelbare spørsmålene til denne oppdagelsen var å undersøke den biologiske og kjemiske naturen til den strukturen.
Disse spørsmålene begynte å bli belyst mellom 1870 og 1900, med eksperimentene til Friedrich Miescher, som introduserer ordet nuklein. Imidlertid endrer Walther Flemming begrepet og bruker Kromatin For å referere til det nukleære stoffet.
Over tid begynner du å ha en dypere kunnskap om genetisk materiale og dens egenskaper. Det er først i 1908 når forskeren fra Italia Pasquale Baccarini bemerker at kromatin ikke er homogen og klarer å visualisere små kropper inne i kjernen.
Typene kromatiner - eukromatin og heterokromatin - ble opprinnelig foreslått av Emil Heitz i 1928. For å etablere denne klassifiseringen var Heitz basert på bruk av farging.
I 1974 foreslo biologen Roger Kornberg en modell for organisering av genetisk materiale i strukturer kjent som nukleosomer, hypotese empirisk bekreftet av Markus nolleksperimenter.
Hva er kromatin?
Kromatinkomponenter: DNA og proteiner
Kromatin er et nukleoprotein dannet av foreningen av genetisk materiale - DNA - med et heterogent sett med protein. Denne assosiasjonen er svært dynamisk og tilegner seg en kompleks tredimensjonal konformasjon som lar den utføre sin regulatoriske og strukturelle funksjoner.
Kan tjene deg: profaseEt av de viktigste kromatinproteinene er histoner, som nesten er i like stor andel som DNA.
Histonas er grunnleggende proteiner, bemerkelsesverdig bevart i hele den evolusjonshistorien til organiske vesener - det vil si at histonene våre ikke varierer mye sammenlignet med andre pattedyr, til og med et annet fjernere dyr fylogenetisk.
Histonens belastning er positiv, slik at de kan samhandle gjennom elektrostatiske krefter med den negative belastningen på fosfatskjelettet som er til stede i DNA. Det er fem typer histoner, nemlig: H1, H2A, H2B, H3 og H4.
Det er også en serie proteiner av en annen karakter enn histoner som deltar i DNA -komprimering.
DNA -komprimering: nukleosomer
Den grunnleggende enheten til kromatin er nukleosomer - repeterende strukturer dannet av DNA og histoner, konformasjon som vi finner gjennom det genetiske materialet.
Den doble DNA -propellen rulles i et åtte histonkompleks kjent som Histonas Octa. Molekylet rulles i omtrent to omganger, etterfulgt av et kort område (mellom 20 og 60 basepar) som skiller nukleosomer fra hverandre.
For å forstå denne organisasjonen, må vi ta hensyn til at DNA -molekylet er ekstremt langt (ca. 2 meter) og må være påmeldt på en ryddig måte å bosette seg i kjernen (hvis diameter er 3 til 10 um). I tillegg må den være tilgjengelig for replikering og transkripsjon.
Dette målet oppnås med forskjellige nivåer av komprimering av DNA, og er den første av dem de nevnte nukleosomene. Disse ligner beretningene til et perlehalskjede. Cirka 150 DNA -basepar rulles hjemme "Konto".
I bakterier er det ingen sanne historier. I kontrast er det en serie proteiner som minner histoner, og det antas at de bidrar til bakteriell DNA -emballasje.
Høyere organisering
Organiseringen av kromatin er ikke begrenset på nivået av nukleosomer. Denne protein- og DNA -assosiasjonen er gruppert i en tykkere struktur på omtrent 30 nm - på grunn av denne tykkelsen kalles det nivået "30 nm fibernivå".
Kromatin organisert i tykkelsen 30 nm er organisert etter tur i form av løkker som strekker seg i et slags protein natur stillas (ikke histoner).
Dette er modellen som for øyeblikket håndteres, selv om eksistensen av mer komplekse komprimeringsmekanismer kan forventes. Den endelige organisasjonen består av kromosomet.
Det kan tjene deg: Cilia: Kjennetegn, struktur, funksjoner og eksemplerFeil i kromatinorganisasjon
Komprimeringen og organiseringen av det genetiske materialet er viktig for flere biologiske funksjoner. Ulike medisinske tilstander har vært assosiert med feil i strukturen til kromatin, inkludert Alpha Talasia knyttet til X-kromosom, Rubinstein-Taybi-syndrom, Coffin-Lowry Syndrome, Rett Syndrome, blant andre.
Typer kromatin

Det er to typer kromatin i cellen, avslørt ved påføring av farging: eukromatin ("true" kromatin) og heterokromatin. I det første tilfellet blir fargingen observert svakt, mens fargingen i det andre er intens.
Denne strukturelle organisasjonen av DNA er unik for eukaryote organismer og er avgjørende for atferden til kromosomer og regulering av genuttrykk.
Hvis vi evaluerer proporsjonene av begge typer kromatiner i en celle som er i grensesnitt, finner vi at omtrent 90% av kromatinet er eukromatin og de resterende 10% tilsvarer heterokromatin. Neste vil vi beskrive hver type i detalj:
Yo. Heterokromatin
Kjennetegn
Hovedforskjellen mellom begge typer kromatin er relatert til graden av komprimering eller "pakking" av molekylet i spesifikke stadier i celledelingen.
Selv om det i grensesnittet ser ut til å være tilfeldig spredt, er det ikke på denne måten.
Det er en betydelig organisasjon på dette stadiet, hvor du kan se en differensiell partisjon av kromosomalt materiale inne i kjernen.
Kromosomene DNA er ikke sammenvevd med DNA -strengen til andre kromosomer, og restene er spesifikke regioner som kalles kromosomale territorier. Denne organisasjonen ser ut til å bidra til genuttrykk.
Heterokromatin er sterkt kondensert, og er lite tilgjengelig for transkripsjonsmaskiner - slik at de ikke blir transkribert. I tillegg er det dårlig med tanke på mengden gener det presenterer.
Typer heterokromatin
Visse heterokromatinregioner er vedvarende gjennom cellelinjer - det vil si, alltid Det vil oppføre seg som heterokromatin. Denne typen heterokromatin er kjent som konstitutiv. Et eksempel på dette er de kondenserte regionene i kromosomene som kalles sentromerer og telomerer.
I kontrast er det heterokromatindeler som kan variere komprimeringsnivået som respons på endringer i utviklingsmønstre eller miljøvariabler.
Takket være de nye studiene blir denne visjonen omformulert, og nå er det bevis på at konstitutiv heterokromatin også er dynamisk og i stand til å svare på stimuli.
Struktur
En av faktorene som bestemmer strukturen til kromatin er de kjemiske modifikasjonene av histoner. Når det gjelder kromatin som er transkripsjonelt inaktivt, viser de hypoacetiserte histoner.
Kan tjene deg: Peroxidases: Struktur, funksjoner og typerNedgangen i mengden av acetylgrupper er assosiert med stillhet av gener, siden den positive belastningen på lisinene ikke vil masca, noe som tillater en sterk elektrostatisk interaksjon mellom DNA og histoner.
Et annet epigenetisk merke er metylering. Siden tilsetning av en metylgruppe ikke modifiserer belastningen på proteinet, er dens konsekvens (aktiverer eller deaktiverer gener) ikke så åpenbar og vil avhenge av regionen til histonet der merkevaren er lokalisert.
Empirisk har det blitt funnet at metylering av H3K4me3 og H3K36me3 er assosiert med genaktivering og de til H3K9me3 og H3K27me3.
Funksjoner
I eksemplet med konstitutiv heterokromatin nevner vi sentromere. Denne kromosomale regionen har en strukturell rolle og bidrar til bevegelse av kromosomer under både mitotiske og meiotiske celledelingshendelser.
Ii. Eukromatin
Kjennetegn
I motsetning til heterokromatin, er eukromatin et mindre kompakt molekyl, så transkripsjonsmaskineriet har enkel tilgang (spesielt til enzymet RNA -polymerase) og kan uttrykkes med aktivt genetiske veier.
Struktur
Strukturen til et nukleosom av et aktivt kromatin fra transkripsjonens synspunkt er preget av acetylerte histoner. Tilstedeværelsen av monometyllisiner er også assosiert med genaktivering.
Tilsetning av en acetylgruppe til dette lysinavfallet fra histonene nøytraliserer den positive belastningen av nevnte aminosyre. Den umiddelbare konsekvensen av denne endringen er reduksjon av elektrostatiske interaksjoner mellom histon og DNA, og genererer et mer slapp kromatin.
Denne strukturelle modifiseringen tillater interaksjonene mellom genetisk materiale med transkripsjonsmaskineriet, som er preget av å være spesielt klumpete.
Funksjoner
Eukromatin omfatter alle gener som er aktive og klarer å oppnås ved transkripsjon -relatert enzymatiske maskiner. Derfor er funksjonene like brede som funksjonene til de involverte genene.
Referanser
- Grewal, s. Yo., & Moazed, d. (2003). Heterokromatin og epigenetisk kontroll av genuttrykk. Vitenskap, 301(5634), 798-802.
- Jost, k. L., Bertulat, f., & Cardoso, m. C. (2012). Heterokromatin og genposisjonering: Innvendig, utenfor, hvilken som helst side?. Kromosom, 121(6), 555-563.
- Lewin, f. (2008). IX -gener. Jones og Bartlett Publisher.
- Tollefsbol, t. ENTEN. (2011). Håndbok for epigenetikk. Akademisk presse.
- Wang, J., Jia, s. T., & Jia, S. (2016). Ny innsikt i reguleringen av heterokromatin. Trender i genetikk: tig, 32(5), 284-294.
- Zhang, p., Torres, k., Liu, x., Liu, c. G., & Pollock, r. OG. (2016). En oversikt over kromatinregulerende proteiner i celler. Nåværende protein og peptidvitenskap, 17(5), 401-410.
- « Ergastiske stoffer klassifisering og biologisk betydning
- Yucca filifera -egenskaper, taksonomi, habitat, bruk »