

Celledelingstyper, prosesser og betydning
- 1348
- 383
- Daniel Skuterud
De Cellular Division Det er prosessen som lar alle levende organismer vokse og reprodusere. I prokaryoter og eukaryoter er resultatet av celledeling datterceller som har samme genetiske informasjon som den opprinnelige cellen. Dette skjer fordi informasjonen i DNA før divisjonen er doblet.
I prokaryoter skjer divisjon av binær fisjon. Genomet til de fleste prokaryoter er et sirkulært DNA -molekyl. Selv om disse organismer ikke har noen kjerne, er DNAet i en komprimert form som kalles nukleoid, som skiller seg fra cytoplasma som omgir det.
Kilde: Retama [CC BY-SA 3.0 (http: // creativecommons.Org/lisenser/by-SA/3.0/]] I eukaryoter skjer divisjon gjennom mitose og meiose. Det eukaryote genomet består av store mengder organisert DNA i kjernen. Denne organisasjonen er basert på DNA -emballasje med proteiner, og danner kromosomer, som inneholder hundrevis eller tusenvis av gener.
De veldig mangfoldige eukaryotene, både encellede og metazoarier, har livssykluser som veksler mitose og meiose. Disse syklusene er de med: a) gammatic meiose (dyr, noen sopp og alger), b) cygotisk meiose (noen sopp og protozoer); og c) veksling mellom gamatisk og cygotisk meiose (planter).
[TOC]
Folkens
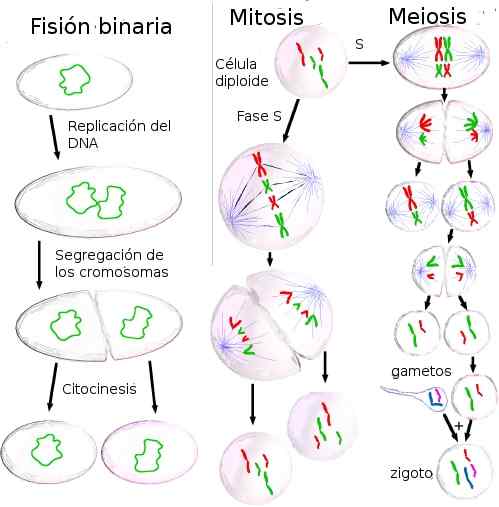
Celledeling kan være ved binær fisjon, mitose eller meiose. Følgende beskriver hver av prosessene som er involvert i denne typen celledeling.
Binær fisjon
 Procariotic fisjon, binær fisjon, er en aseksuell reproduksjonsform.
Procariotic fisjon, binær fisjon, er en aseksuell reproduksjonsform. Binær fisjon består av delingen av cellen som gir opphav til to datterceller, hver med en identisk kopi av det originale celle -DNA.
Før delingen av den prokaryote cellen finner replikasjonen av DNA sted, som begynner på et bestemt sted for dobbeltkjeden DNA, kalt replikasjonens opprinnelse. Replikasjonsenzymer beveger seg mot begge retningene for opprinnelsen, og produserer en kopi av hver av Double Chain DNA -kjedene.
Etter DNA -replikering forlenges cellen og DNAet skilles inn i cellen. Umiddelbart begynner en ny plasmamembran å vokse midt i cellen, og danner et septum.
Denne prosessen tilrettelegges av FTSZ -proteinet, som evolusjonært er veldig bevart i prokaryoter, inkludert archaea. Endelig er cellen delt.
Cellesyklus og mitose
Stadiene som en eukaryotisk celle fra to påfølgende celledelinger er kjent som en cellesyklus. Varigheten av cellesyklusen varierer fra noen minutter til måneder, avhengig av celletype.
Cellesyklusen er delt inn i to trinn, nemlig fase m og grensesnitt. To prosesser, kalt mitose og cytokinesis forekommer i M -fasen. Mitose består av atomavdeling. Det samme antallet og typene kromosomer som er til stede i den opprinnelige kjernen finnes i barnekjernene. Somatiske celler av flercellede organismer er delt av mitose.
Cytokinesis består av cytoplasma -divisjonen for å danne datterceller.
Grensesnittet har tre faser: 1) G1, cellene vokser og bruker mesteparten av tiden sin i denne fasen; 2) S, genom duplisering; og 3) G2, replikering av mitokondrier og andre organeller, kondensering av kromosomer og mikrotubuli -montering, blant andre hendelser.
Mitosetrinn
Mitose begynner med slutten av G2 -fasen, og er delt inn i fem faser: Profase, Promise, Metafase, Anafase og Telophase. De skjer alle kontinuerlig.
Kan tjene deg: Cellulær kommunikasjon: Typer, betydning, eksemplerProfase
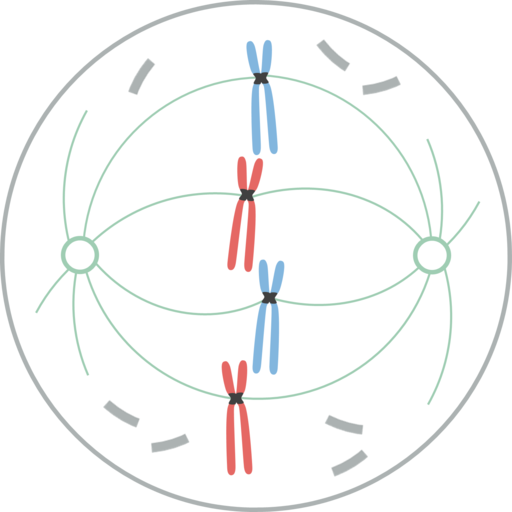 Profase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons
Profase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons På dette stadiet er monteringen av det mitotiske spindelen, eller mitotisk apparat, hovedbegivenheten. Profase begynner med kromatinkomprimering, og danner kromosomer.
Hvert kromosom har et par søstre kromater, med identisk DNA, som er nært knyttet sammen i nærheten av deres sentromerer. Proteinkomplekser kalt kohesiner deltar i denne foreningen.
Hver sentromere er koblet til en cinetocoro, som er et proteinkompleks som binder seg til mikrotubuli. Disse mikrotubuloene lar hver kopi av kromosomene tildeles dattercellene. Microtubules radian fra hver ende av cellen og danner mitotiske apparater.
I dyreceller, før profase, oppstår duplisering av sentrering, som er det viktigste organiseringssenteret for mikrotubulene og stedet der foreldrene og Son Centrioles er lokalisert. Hver sentrering når motsatt pol av cellen, og etablerer en mikrotubulosbro mellom dem kalt mitotisk enhet.
I de nyeste evolusjonsplantene, i motsetning til dyreceller, er det ingen sentre, og opprinnelsen til mikrotubuli er ikke klart. I fotosyntetiske celler med eldre evolusjonær opprinnelse, for eksempel grønne alger, er det sentre.
Love
Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] Mitose skal garantere segregeringen av kromosomer og fordelingen av kjernefysisk konvolutt til kjernefysisk pore og nukleoluskompleks. Avhengig av om kjernefysisk innpakning (IN) forsvinner eller ikke, og graden av denstegrering av IN, varierer mitosen fra lukket til helt åpen.
For eksempel i S. Cerevisae Mitose er lukket, i TIL. Nidulans Det er semi -åpent, og hos mennesker er det åpent.
I lukket myitose er spindelens polare kropper inne. Cytoplasmatiske mikrotubuli samhandler med cellebarken, og med kromosomene knadere.
I semi -Absert myitose, fordi IN -er delvis desensamblada, invaderes det kjernefysiske rommet av mikrotubulene som er kjernet fra sentrene og gjennom to åpninger av IN, dannende laget omgitt av in.
Ved åpen mitose forekommer den komplette de -slasamblagen i, det mitotiske apparatet er ferdig og kromosomer begynner å bli forskjøvet mot midten av cellen.
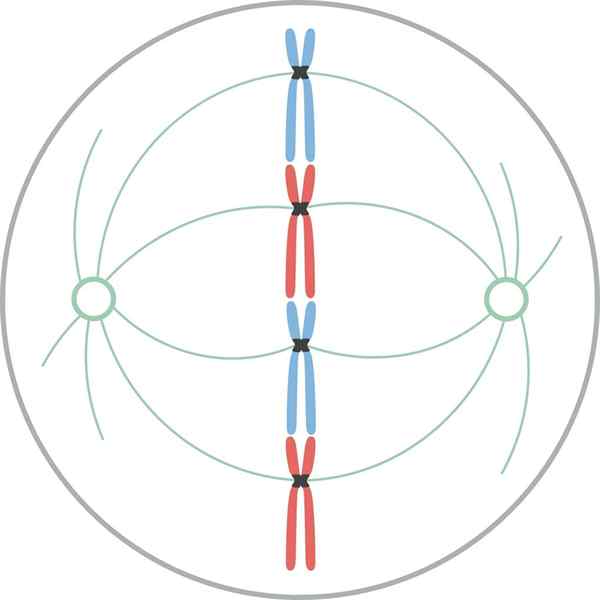
Metafase
Kromosomer justert i ekvatorialplaten til cellen under mytisk metafase I metafase er kromosomer justert i cellen til cellen. Det imaginære planet vinkelrett på spindelaksen, som passerer gjennom den indre omkrets av cellen, kalles metafasetretten.
I pattedyrceller er det mitotiske apparatet organisert i en sentral mitotisk spindel og et par osterer. Den mitotiske spindelen består av et symmetrisk bilateralt bunt med mikrotubuli som er delt inn i cellen til cellen, og danner to motsatte halvdeler. Ásteres består av en gruppe mikrotubuli i hver spindelstang.
I det mitotiske apparatet er det tre grupper av mikrotubuli: 1) Astral, som danner Rster, starter fra sentrering og stråler mot cellebarken; 2) av cinetocoro, som binder seg til kromosomene gjennom cinetocoro; og 3) polar, som interdign med mikrotubuli av den motsatte polen.
I alle mikrotubuli beskrevet ovenfor er endene (-) orientert mot sentrene.
I planteceller, hvis det ikke er noen sentrering, er spindelen lik den for dyreceller. Spindelen består av to halvdeler med motsatt polaritet. Endene (+) finnes i ekvatorialplaten.
Det kan tjene deg: Podocytter: Karakteristikker, histologi og funksjonerAnafase
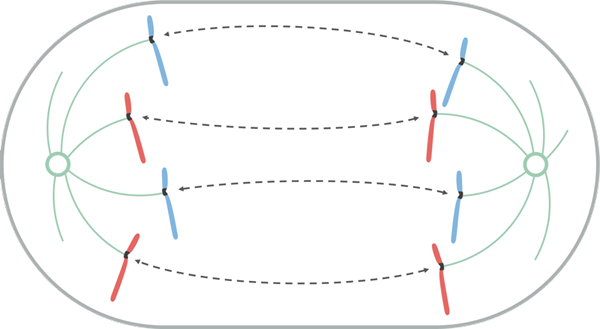 Kilde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons
Kilde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons Anafase er delt tidlig og sent. I tidlig anafase finner separasjonen av søsterkromatider sted.
Denne separasjonen oppstår fordi proteinene som opprettholder unionen er kledd og fordi det er en forkortelse av kynetokoro -mikrotubuli. Når paret med søsterkromatider er separert, kalles de kromosomer.
Under forskyvningen av kromosomene mot polene beveger cinetocoro seg langs mikrotubulen til den samme cynetocoro som dens ende (+) dissosiater. På grunn av dette er bevegelsen av kromosomer under mitose en passiv prosess som ikke trenger motoriske proteiner.
I den sene anafasen skjer en større separasjon av polene. Et KRP -protein, knyttet til det ekstreme (+) av polare mikrotubuli, i overlappingsområdet med det samme, marsjerer mot slutten (+) av en tilstøtende polar mikrotubulus antiparallell. Dermed skyver KRP den tilstøtende polare mikrotubulen mot slutten (-).
I planteceller, etter separasjon av kromosom. Denne strukturen tillater begynnelsen på det cytocinetiske apparatet, kalt Framoplasto.
Telofase
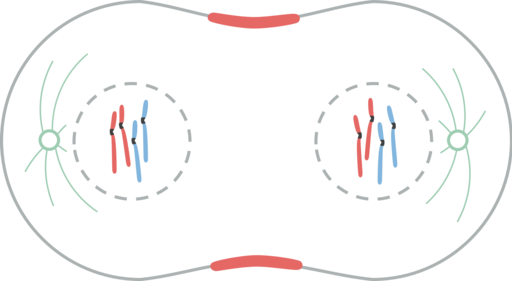 Telofase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)]
Telofase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] I telofasen skjer flere hendelser. Kromosomer når poler. Cinetocoro forsvinner. Polare mikrotubuli fortsetter å forlenge seg selv, og forbereder cellen for cytokinesis. Atomkonvolutten dannes igjen fra fragmenter av mors innpakning. Kjernen dukker opp igjen. Kromosomer blir misforstått.
Cytokinesis

Citocinesis er fasen av cellesyklusen som cellen er delt. I dyreceller oppstår cytosinesis ved hjelp av en aktinfilament innsnevringsstropp. Disse filamentene glir over hverandre, diameteren på beltet avtar, og en spor av Clivaje dannes rundt celleomkretsen.
Fordi innsnevringen fortsetter, blir sporet utdypet og en intercellulær bro dannes, som inneholder gjennomsnittlig kropp. I den sentrale regionen av den intercellulære broen er bjelkene i mikrotubulene, som er dekket av en elektrotensmatrise.
Rupturen av den intercellulære broen mellom post-mitotiske søsterceller finner sted gjennom abscission. Det er tre typer abscisjon: 1) mekanisk bruddmekanisme; 2) arkiveringsmekanisme etter interne vesikler; 3) Plasmamembranens innsnevring for fisjon.
I planteceller er membrankomponenter samlet inni og celleplaketten dannes. Denne platen vokser til overflaten av plasmamembranen, smelter sammen med den og deler cellen i to. Deretter blir cellulosen avsatt på den nye plasmamembranen og danner den nye celleveggen.
Meiose
Meiose er en type celledeling som reduserer antall kromosomer i to. Dermed er en diploid celle delt inn i fire haploide datterceller. Meiose skjer i spirende celler og gir opphav til gameter.
Stadiene av meiose består av to inndelinger av kjernen og cytoplasma, nemlig meiose I og meiose II. Under meiose I skiller medlemmene av hvert par homologe kromosomer. Under meiose II produseres søsterkromatider separere og fire haploide celler produseres.
Hvert trinn med mitose er delt inn i profase, lovet, metafase, anafase og telofase.
Kan tjene deg: bakteriecellevegg: egenskaper, biosyntese, funksjonerMeiose i
- Profase i. Kromosomene kondenserer og spindelen begynner å danne seg. DNA er doblet. Hvert kromosom består av søsterkromatider, sammen med sentromere. Homologe kromosomer vises under synapse, noe som tillater kryss -kobling, som er nøkkelen til å produsere forskjellige gameter.
- Metafase i. Paret av homologe kromosomer er rettet langs metafaseplaten. Chiasma hjelper med å holde det vedlagte paret. Mikrotubuli av cinetocoro av hver pol blir med en sentromere av et homologt kromosom.
- Anafase i. Mikrotubuli av cinetocoro er forkortet og de homologe parene er atskilt. En duplikat kollega går til en cellestang, mens den andre duplikatmotparten går til den andre siden av polet.
- Telofase i. Separate homologer danner en gruppe i hver cellestang. Atomkonvolutten er igjen. Citocinesis skjer. De resulterende cellene har halvparten av antall originale cellekromosomer.
Meiose II
- Profase II. En ny spindel dannes i hver celle og cellemembranen forsvinner.
- Metafase II. Spindelformasjonen er fullført. Kromosomene har søstr. Mikrotubulene i Cinetocoro som starter fra motsatte poler binder seg til sentromereren.
- Anafase II. Mikrotubrene er forkortet, sentromererene er delt, søsterkromatidene skiller seg og beveger seg mot motsatte poler.
- Telofase II. Nukleær innpakning rundt fire grupper av kromosomer dannes: Fire haploide celler dannes.
Betydning
Gjennom noen eksempler er viktigheten av de forskjellige typer celledeling illustrert.
- Mitose. Cellesyklusen har irreversible punkter (DNA -replikasjon, separasjon av søsterkromatider) og kontrollpunkter (G1/s). P53 -protein er nøkkelen til G1 -kontrollpunktet. Dette proteinet oppdager DNA -skade, stopper celledelingen og stimulerer aktiviteten til enzymer som reparerer skaden.
I mer enn 50% av kreft i mennesker har p53 -proteinet mutasjoner som annullerer sin evne til å sette spesifikke DNA -sekvenser. P53 -mutasjoner kan være forårsaket av kreftfremkallende.
- Meiose. Er assosiert med seksuell reproduksjon. Fra det evolusjonære synspunktet antas det at seksuell reproduksjon oppsto som en prosess for å reparere DNA. Dermed kan skaden som produseres i et kromosom repareres basert på informasjonen til det homologe kromosomet.
Det antas at den diploide staten var forbigående i gamle organismer, men at den begynte å ha mer relevans etter hvert som genomet ble større. I disse organismer har seksuell reproduksjon komplementering, DNA -reparasjon og genetisk variasjon.
Referanser
- Alberts, f., Johnson, a., Lewis, J., og kål. 2007. Biologi av cellemolekylæren. Garland Science, New York.
- Bernstein, h., Byers, g.S., Michod, r.OG. 1981. Evolusjon av seksuell reproduksjon: Viktigheten av DNA -reparasjon, komplementering og variasjon. American Naturalist, 117, 537-549.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, p., Baltimore, d., Darnell, J. 2003. Cellulær og molekylær biologi. Pan -American Medica Redaksjon, Buenos Aires.
- Raven, s.H., Johnson, g.B., Losos, J.B., Singer, s.R. 2005 Biologi. Høyere utdanning, Boston.
- Solomon, f.M., Berg, l.R., Martin, d.W. 2008. biologi. Thomson, USA.
- « Teporingo -egenskaper, fare for utryddelse, habitat
- Ricinus kommuneegenskaper, habitat, toksisitet, bruker »