

DNA -emballasje
- 1471
- 174
- Markus Fredriksen
DNA -emballasjeopplegg. Kilde: Thomas Splettstoesser (www.Scistyle.com), CC av 4.0, Wikimedia Commons Hva er DNA -emballasje?
Han DNA -emballasje Det er et begrep som definerer den kontrollerte komprimeringen av DNA inne i cellular. DNA er et ekstremt langt molekyl som i tillegg alltid samhandler med et stort utvalg av forskjellige proteiner (nukleoproteiner). Med dem dannes kromatinet, som er stoffet som komponerer kromosomene.
For behandling, arv og kontroll av uttrykket av genene som DNA vedtar en bestemt romlig organisasjon. Cellen oppnår den som strengt kontrollerer DNA -emballasje på forskjellige komprimeringsnivåer.
Virus har forskjellige emballasjestrategier for nukleinsyrene. En av favorittene er dannelsen av kompakte spiraler. Det kan sies at virus er nukleinsyrer pakket i proteinene som dekker dem, beskytter og mobiliserer.
I prokaryoter er DNA assosiert med proteiner som bestemmer dannelsen av komplekse bånd i en struktur som kalles nukleoid. Det maksimale komprimeringsnivået av DNA i en eukaryotisk celle, derimot, er det mitotiske eller meiotiske kromosomet.
DNA -struktur
DNAet består av to antipaallale bånd (de løper i motsatt retning) som danner en dobbel propell. Hver av dem presenterer et skjelett av fosfodisterbindinger som sukker knyttet til nitrogenbaser er sammen med.
Inne.
I et molekyl som dette viser viktigste koblingsvinkler gratis rotasjon. Nitrogen-sukkerbindinger, sukkergruppefosfat og fosfodiésterbinding er fleksible.
Dette tillater DNA, sett på som en fleksibel stang, viser en viss kapasitet til å bøye og rulle. Denne fleksibiliteten lar deg ta i bruk komplekse lokale strukturer, og danne korte, middels og langdistanseinteraksjonsbånd.
Kan tjene deg: Hva er den kromosomale arveteorien? (Sutton og Morgan)Denne fleksibiliteten forklarer også hvordan 2 meter DNA kan opprettholdes i hver diploide celle av et menneske. I en gamete (haploidcelle) ville det være en meter DNA.
Bakteriekjernen
Selv om det ikke er en urokkelig regel, eksisterer bakteriekromosomet som et enkelt dobbeltbånd over -gleaming DNA -molekyl.
Den doble propellen handler mer om seg selv (mer enn 10 bp ved retur) og produserer dermed en viss komprimering. Lokale knuter genereres også takket være manipulasjoner som er enzimatisk kontrollert.
I tillegg er det DNA -sekvenser som lar domener danne seg i store bånd. Strukturen som følge av super -collapse og bånd som er bestilt, kalles nukleoid.
Disse opplever dynamiske endringer takket være noen proteiner som gir en viss strukturell stabilitet til det komprimerte kromosomet. Graden av komprimering i bakterier og buer er så effektiv at det kan være mer enn en nukleoidkromosom.
Den kompakte nukleoid DNA -prokaryoter minst 1.000 ganger. Den nukleoide topologiske strukturen er en grunnleggende del av reguleringen av genene som kromosomet bærer. Det vil si struktur og funksjon utgjør samme enhet.
Eukaryote kromosomkomprimeringsnivåer
DNA i den eukaryote kjernen er ikke naken. Samhandle med mange proteiner, hvorav de viktigste er histoner. Histonas er positivt lastede små proteiner som binder seg til DNA på en bestemt måte.
I kjernen er det vi observerer et komplekst DNA: histoner, som vi kaller kromatin. Svært kondensert kromatin, som vanligvis ikke er uttrykt, er heterokromatin. Tvert imot, den minst komprimerte (mer åpne), eller eukromatin, er kromatin med gener som kommer til uttrykk.
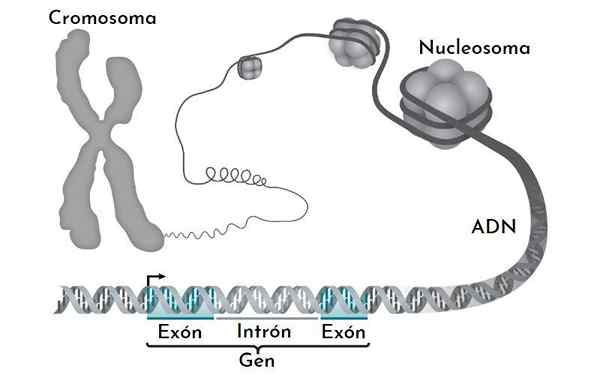
Kan tjene deg: ren linjeKromatin har flere komprimeringsnivåer. Den mest elementære er nukleosomet. De blir fulgt av magnetfiber og grensesnittkromatinbånd. Bare når et kromosom er delt er at maksimale komprimeringsnivåer vises.
Nukleosomet
Nukleosom er den grunnleggende enheten i kromatinorganisasjonen. Hvert nukleosom dannes av en okta av histoner som danner en slags trommel.
Oktameren er dannet av to eksemplarer av hver av H2A, H2B, H3 og H4 histoner. Rundt dem gir DNA nesten 1.7 svinger. Det blir fulgt av en fri DNA -fraksjon kalt linker, 20 bp, assosiert med histon H1, og deretter en annen nukleosom. Mengden DNA i ett nukleosom og den som blir med det er omtrent 166 basepar.
Dette pakketrinnet med kompakt DNA molekylet omtrent 7 ganger. Det vil si at vi passerer en meter til litt over 14 cm DNA.
Denne emballasjen er mulig fordi positive histoner kansellerer den negative belastningen av DNA, og den påfølgende elektrostatiske selvtallet. Den andre grunnen er at DNA kan brettes på en slik måte at det kan sirkle Histone Octamer.
30 nm fiber
Regnskapsfiberen i et halskjede som danner mange påfølgende nukleosomer er i tillegg melder seg i en mer komprimert struktur.
Selv om vi ikke er sikre på hvilken struktur som virkelig vedtar, vet vi at den når en tykkelse på omtrent 30 nm. Dette er den så -kalt 30 nm fiberen, og for dens dannelse og stabilitet er histonen H1 grunnleggende.
30 nm fiber er den grunnleggende strukturelle enheten til heterokromatin. Det av slappe nukleosomer, eukromatin.
Bånd og svinger
30 nm fiber er imidlertid ikke helt lineær. Tvert imot danner det bånd på omtrent 300 nm lang, på en svingete måte, på en litt kjent proteinmatrise.
Kan tjene deg: rettsmedisinsk genetikk: historie, studieobjekt, metodikkDisse båndene på en proteinmatrise danner en mer kompakt kromatinfiber 250 nm i diameter. Til slutt samsvarer de som en enkel 700 nm tykk propell, og gir opphav til en av søsterkromatider i et mitotisk kromosom.
Til slutt komprimerer DNA i nukleær kromatin ca.000 ganger på cellekromosomet i divisjon. I grensesnittkjernen er komprimeringen også høy, siden den er omtrent 1.000 ganger sammenlignet med "lineær" DNA.
Den meiotiske komprimeringen av DNA
I utviklingsbiologiens verden sies det at gameteogenese tilbakestiller epigenom. Det vil si at det sletter DNA -merkene at livet til de som ga opphav til gameto produserte eller erfarne.
Disse merkene inkluderer metylering av DNA og de kovalente modifikasjonene av histonene (histonkode). Men ikke alt epigenom tilbakestilles. Det som gjenstår med merkevarer vil være ansvarlig for det mors genetiske avtrykket.
Den implisitte tilbakestillingen til gametoogenese er lettere å se den i sæd. I sæd er DNA ikke pakket med histoner. Derfor er informasjonen som er knyttet til dens modifikasjoner i det produserende organet, generelt ikke arvet.
I sæd pakkes DNA takket være interaksjon med unionsproteiner som ikke er spesifikke for DNA, kalt protaminer. Disse proteinene danner disulfidbroer med hverandre, og bidrar dermed til å danne overfylte lag som ikke frastøter elektrostatisk.
Referanser
- DNA -emballasje: Nukleosomer og kromatin. Hentet fra naturen.com.