

Erythropoietin (EPO) egenskaper, produksjon, funksjoner
- 1574
- 131
- Daniel Skuterud
De erytropoietin, hemopoietin eller Epo Det er et glykoprotein med hormonelle funksjoner (cytoquin) som har ansvar for spredning, differensiering og overlevelseskontroll av stamfadercellene til erytrocytter eller røde blodlegemer i benmargen, det vil si erytropoiesisisisisisisisisisisisisisisisis.
Dette proteinet er en av de forskjellige vekstfaktorene som kontrollerer de hematopoietiske prosessene som de dannes, fra en liten gruppe pluripotensielle stamceller, cellene som finnes i blodet: både erytrocytter og hvite blodlegemer og lymfocytter. Det vil si myeloide og lymfoide avstamningsceller.
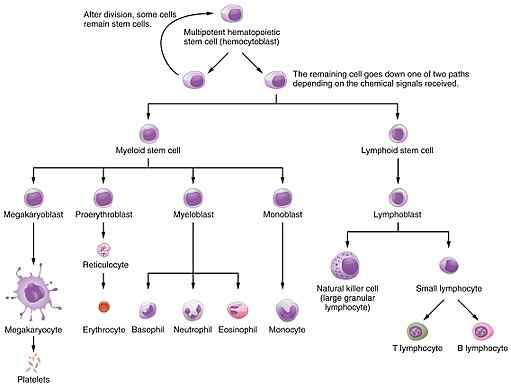
Ordning som representerer hemopoiesis, hvor prosessen med dannelse av erytrocytter eller erytropoiesis er inkludert, der erytropoietin handlinger (kilde: OpenStax College [CC av 3.0 (https: // creativecommons.Org/lisenser/av/3.0)] via Wikimedia Commons) Dets betydning ligger i den funksjonelle viktigheten som celler har som hjelper til.
Erythropoietin var den første vekstfaktoren som ble klonet (i 1985), og for tiden er dens administrasjon for den vellykkede anemibehandlingen produsert av nyresvikt godkjent av American Drug and Food Administration (FDA) (FDA).
Forestillingen om at erytropoiesis styres av en humoral faktor (oppløselig faktor til stede i sirkulasjonen) ble foreslått for mer enn 100 år siden av Carnot og DeFlandre når de studerte de positive effektene på økningen i prosentene av røde celler hos kaniner behandlet med serum av anemisk dyr.
Imidlertid var det først i 1948 da Bonsidorff og Jalavist.
[TOC]
Kjennetegn
Erythropoietin er et protein fra glykoproteinfamilien. Den er stabil for syre pH og har omtrent 34 kDa molekylvekt.
Det har omtrent 193 aminosyrer, som inkluderer et hydrofobt N-terminalt område på 27 avfall, som elimineres ved co-translasjonsbehandling; Og en argininrest i posisjon 166 som også går tapt, så proteinet i sirkulasjonen har 165 aminosyrer.
I sin struktur kan dannelsen av to disulfur-broer mellom cysteinavfallet som er til stede i posisjoner 7-161 og 29-33, sees, som er knyttet til deres drift. Det er mer eller mindre enn 50% av alfa -propellene, som tilsynelatende deltar i dannelsen av en region eller en kuleform.
Den har 40% karbohydrater, representert med tre kjeder av oligosakkarider N-enheter til forskjellige asparaginsyre (ASP) rester, og en kjede O-forenet til en serinrest (SER). Disse oligosakkaridene består hovedsakelig.
Epo karbohydratregionen oppfyller flere roller:
Kan tjene deg: Flora og fauna av Coahuila- Det er viktig for biologisk aktivitet.
- Det beskytter mot nedbrytning eller skade forårsaket av oksygenfrie radikaler.
- Oligosakkaridekjeder er nødvendig for moden proteinsekresjon.
Hos mennesker er genet som koder for dette proteinet lokalisert midt i den lange armen til kromosom 7, i Q11-Q22-regionen; Det er i en enkelt kopi i en 5 -region.4KB og har fem eksoner og fire introner. Homologistudier indikerer at sekvensen deres deler 92% identitet med andre primater og 80% med den for noen gnagere.
Produksjon
I fosteret
Under fosterutvikling produseres erytropoietin hovedsakelig i leveren, men det har blitt bestemt at i løpet av samme stadium er genet som koder for dette hormonet også uttrykt rikelig i den gjennomsnittlige regionen av nyrens nefroner.
I voksen
Etter fødselen, i hva alle postnatale stadier kan vurderes, produseres hormonet i det vesentlige i nyrene. Spesielt av cortex -celler og overflaten av nyrekorpuscles.
Leveren deltar også i produksjonen av erytropoietin i postnatal stadiene, der mer eller mindre 20% av den totale EPO i sirkulasjonen skilles ut.
Andre "ekstra nyre" organer der erytropoietinproduksjon er blitt påvist inkluderer perifere endotelceller, vaskulære glatte muskelceller og insulinproduserende celler.
Det er også kjent at det i sentralnervesystemet er noen EPO -sekresjonssentre, inkludert hippocampus, cortex, cerebrale endotelceller og astrocytter.
Erythropoietin produksjonsregulering
Erytropoietinproduksjon kontrolleres ikke direkte av mengden røde blodlegemer i blodet, men av tilførsel av oksygen i vev. En oksygenmangel i vevene stimulerer produksjonen av EPO og dets reseptorer i leveren og nyrene.
Denne aktiveringen av genuttrykk mediert av hypoksi er produktet av aktiveringen av ruten til en familie av transkripsjonsfaktorer kjent som en faktor 1 inducerbar ved hypoksi (HIF-1, av engelsk Hypoksi-indusibel faktor 1).
Hypoksi induserer da dannelsen av mange proteinkomplekser som oppfyller forskjellige funksjoner i aktiveringen av erytropoietinuttrykk, og som er direkte eller indirekte forbundet med faktorer som oversetter aktiveringssignalet til promoteren av EPO -genet, stimulerer transkripsjonen.
Andre stressende faktorer som hypoglykemi (lavt blodsukkerkonsentrasjon), intracellulært kalsium øker eller tilstedeværelsen av reaktive oksygenarter utløser også HIF-1-ruten.
Kan tjene deg: homopolysakkarider: egenskaper, struktur, funksjoner, eksemplerVirkningsmekanismen
Virkningsmekanismen til erytropoietin er ganske kompleks og avhenger hovedsakelig av dens evne til å stimulere forskjellige signaliserende fossefall involvert i celleproliferasjon, som på sin side er relatert til aktivering av andre faktorer og hormoner.
I menneskekroppen til en sunn voksen er det en balanse mellom produksjon og ødeleggelse av røde blodlegemer eller erytrocytter, og EPO deltar i opprettholdelsen av denne balansen gjennom erstatning av erytrocyttene som forsvinner.
Når mengden oksygen tilgjengelig i vevene er veldig lav, øker ekspresjonen av kodingsgenet for erytropoietin i nyrene og leveren. Stimulansen kan også oppstå på grunn av store høyder, hemolyse, tilstander med alvorlig anemi, blødninger eller langvarig eksponering for karbonmonoksid.
Disse forholdene genererer en tilstand av hypoksi, som får sekresjonen av EPO til å øke, det er et større antall røde celler og fraksjonen av retikulocytter i sirkulasjon, som er en av stamfadercellene til erytrocytter, øker også også.
Hvem EPO handler på?
I erytropoiesis deltar EPO hovedsakelig i spredning og differensiering av stamfadercellene som er begått i avstamningen til de røde blodcellene (erytrocyttforeldre), men aktiverer også mitose i proeritroblastene og i basofilen erythroblasts, og også avslappet av løslatelsen og i basofiliske erythroblasts, og også akselererer og også akselererer og også akselererer og i de basofiliske erythroblasts, og Benmargs retikulocytter.
Det første nivået som proteinet fungerer, er i forebygging av programmert celledød (apoptose) av forløpercellene dannet i benmargen, noe som oppnår ved hemmende interaksjon med faktorene som er involvert i denne prosessen.
Hvordan fungerer det?
Celler som reagerer på erytropoietin har en spesifikk mottaker for denne kjent som en erytropoietin eller EPOR. Når proteinet danner et kompleks med mottakeren, overføres signalet til innsiden av cellen: mot kjernen.
Det første trinnet for signaloverføring er en konformasjonsendring som oppstår etter foreningen av proteinet med mottakeren, som samtidig er sammen med andre mottakende molekyler som er aktivert. Blant dem er Janus-Pirosina Kinase 2 (Jack-2).
Blant noen av rutene som er aktivert nedstrøms, etter Jack-2 medierer fosforylering av epokreseptorens tyrosinavfall, er ruten til kartkinasene og Quinasa C-proteinet, som aktiverer transkripsjonsfaktorer som øker uttrykket av spesifikke gener.
Det kan tjene deg: paraseksuell reproduksjon: Moneras, Protister og soppFunksjoner
Som mange hormonelle faktorer i organismer, er erytropoietin ikke begrenset til en enkelt funksjon. Dette har blitt belyst gjennom en rekke undersøkelser.
I tillegg til å fungere som en sprednings- og differensieringsfaktor for erytrocytter, som er essensielle for gasetransport gjennom blodomløpet, ser det ut til at erytropoietin oppfyller noen tilleggsfunksjoner, ikke nødvendigvis relatert til cellulær spredning og differensiering cellulær.
I skadeforebygging
Studier har antydet at EPO forhindrer cellulære lesjoner, og selv om deres virkningsmekanismer ikke er kjent nøyaktig, antas det at de apoptotiske prosessene produsert av redusert eller fraværende oksygenspenning kan forhindre, eksitere toksisitet og eksponering for frie radikaler.
I apoptose
Deltakelsen i forebygging av apoptose har blitt studert ved interaksjon med bestemmende faktorer i signalfossene: Janus-Marosina kinase 2 (JAK2), Caspasa 9, Caspasa 1 og Caspasa 3, Glykogensyntasa Volc apoptotiske proteaser 1 (APAF-1) og andre.
Funksjoner i andre systemer
Delta i hemming av cellebetennelse ved å hemme noen proinflammatoriske cytokiner som interleukin 6 (IL-6), alfa-tumor nekrose faktor (TNF-a) og det kjemiske proteinet av monocytter 1.
I det vaskulære systemet er det påvist at det samarbeider om å opprettholde dets integritet og i dannelsen av nye kapillærer fra eksisterende fartøyer i områder uten vaskulatur (angiogenese). I tillegg forhindrer det permeabiliteten til den hematoencephaliske barrieren under skader.
Det antas at det stimulerer postnatal neovaskularisering ved å øke mobiliseringen av stamceller fra benmargen til resten av kroppen.
Det har en viktig rolle i utviklingen av progenitor nevrale celler gjennom aktivering av KB -kjernefaktoren, som fremmer produksjonen av nervestamceller.
EPO handler i konsert med andre cytokiner, og har en "modulerende" funksjon i kontrollen av sprednings- og differensieringsveiene til megacariocytter og granulocytter-monocyttene.
Referanser
- Despoulos, a., & Silbernagl, s. (2003). Atlas of Physiology Color (5. utg.). New York: Thieme.
- Jelkmann, w. (1992). Erytropietin: struktur, kontroll av produksjon og funksjon. Fysiologiske anmeldelser, 72(2), 449-489.
- Jelkmann, w. (2004). Molekylær biologi av erytropoietin. Indremedisin, 43(8), 649-659.
- Jelkmann, w. (2011). Regulering av erytropietinproduksjon. J. Fysiol., 6, 1251-1258.
- Lacombe, ca., & Mayeux, P. (1998). Biologi av erytropietin. Haematologica, 83, 724-732.
- Maiese, k., Li, f., & Zhong, z. (2005). Nye veier for utforskning for erytropietin. Jama, 293(1), 1-6.
- « Selenary acid (H2SO3) egenskaper, risikoer og bruk
- Oxihemoglobin -egenskaper, struktur og krysskurve »