

Flagelina -struktur og funksjoner
- 1827
- 564
- Mathias Aas
De Flagelina Det er et protein av glødetråden, som er en struktur som er en del av svøpet av bakterier. De aller fleste bakterier har bare en type svøpe. Noen har imidlertid mer enn to.
Molekylstørrelsen på dette proteinet varierer mellom 30 kDa og 60 kDa. For eksempel i enterobakterier er molekylær størrelse stor, mens i visse sweetacuícolas -bakterier er den liten.
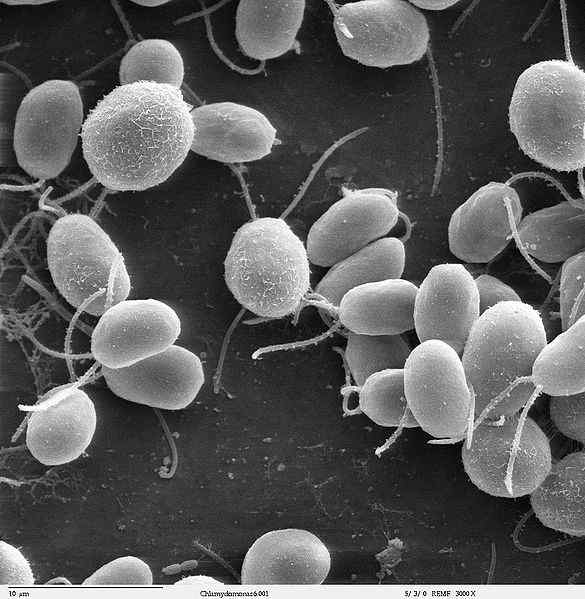
Kilde: Dartmouth Electron Microscope Facility, Dartmouth College [Public Domain] Flagelina er en virulensfaktor som tillater vedheft og invasjon å være vert for celler. I tillegg er det en kraftig aktivator av mange typer celler involvert i den medfødte og adaptive immunresponsen.
[TOC]
Flagelo og mobilitet ultrastruktur
Svøpet er forankret til celleoverflaten. Den består av tre deler: 1) glødetråden, som strekker seg fra overflaten av cellen og er en hul og stiv sylindrisk struktur; 2) basallegemet, som er innebygd i veggene i vegg- og cellemembranen, og danner flere ringer; og 3) kroken, en buet kort struktur som forbinder basallegemet til glødetråden.
Basallegemet er den mest komplekse delen av svøpet. I gram negative bakterier har den fire ringer koblet til en sentral kolonne. I de positive gramene har den to ringer. Rotasjonsbevegelsen av svøpet skjer i basallegemet.
Plasseringen av flagellaen på overflaten av bakteriene varierer mye mellom organismer, og kan være: 1) monric, med bare en svøpe; 2) polar, med to eller flere; eller 3) Peritrico, med mange side flagella. Det er også endoflagelos, som i Spirochetes, som ligger i perplapsmisk rom.
Helicobacter pylori Det er veldig mobil fordi den har seks til åtte unipolare svindel. En pH -gradient gjennom slim tillater H. Pylori Orientere og etablere i et område ved siden av epitelceller. Pseudomonas Den har en polar svøpe, som viser kjemiotaks for sukker og er assosiert med virulens.
Flageline -struktur
Et trekk ved skravlende proteinsekvens er at dens N-terminale og C-terminale regioner er sterkt bevart, mens den sentrale regionen er svært varierende mellom arter og underarter av samme slekt. Denne hypervariabiliteten er ansvarlig for hundrevis av serotyper av Salmonella spp.
Kan tjene deg: Serratia MarcescensFlagelina -molekyler samhandler med hverandre gjennom terminalregionene og polymeriserer og danner et glødetråd. I dette er terminalregionene i det indre av den sylindriske strukturen i glødetråden, mens den sentrale blir utsatt for.
I motsetning til tubulinfilamenter som er depolimert i fravær av salter, er bakterier veldig stabile i vann. Omtrent 20.000 tubulin -underenheter danner et glødetråd.
I glødetråden til H. Pylori og Pseudomonas aeruginosa To typer svøpe og flab er polymerisert, kodet av Flic -genet. Flaa er heterogene og er delt inn i flere undergrupper, med molekylære masser som varierer mellom 45 og 52 kDa. Flab er homogen med en molekylmasse på 53 kDa.
Ofte er flagelin lysinrester metylert. I tillegg er det andre modifikasjoner som Flaa glysilasjon og fosforylering av flab tyrosinavfall, hvis funksjoner er henholdsvis virulens og eksportsignal.
FLAGERY FILAMENT Vekst i bakterier
Bakterienes svøpe kan oppleves eksperimentelt, og være mulig for å studere dens regenerering. Flagelina -underenheter transporteres gjennom den indre regionen i denne strukturen. Når de når slutten, tilsetter underenhetene spontant ved hjelp av et protein ("CAP -protein") kalt HAP2 eller FLOID.
Syntesen av glødetråden finner sted gjennom sin egen forsamling; Det vil si at polymerisasjonen av svøpet ikke krever enzymer eller faktorer.
Informasjonen for montering av glødetråden finnes i selve underenheten. Dermed polymeriserer flagelina seg elleve protofilamenter, som danner en komplett en.
Flagelina syntese av P. Aeruginosa og Proteus mirabilis Det blir hemmet av antibiotika som erytromycin, klaritromycin og azitromycin.
Flagelina som en immunsystemaktivator
De første studiene viste at svøpet, ved subnomolare konsentrasjoner, fra Salmonella, Det er en kraftig cytokininduktor i en promokital cellelinje.
Kan tjene deg: ZoosporesDeretter ble det vist at induksjon av den proinflammatoriske responsen innebærer en interaksjon mellom svøpe og overflateseptorene til de medfødte immunsystemcellene.
Overflatreseptorene som interagerer med flagelin er de av Toll-5 (TLR5) type. Deretter viste studier med rekombinant svøpe at når den manglet den hypervariable regionen, var det ikke i stand til å indusere en immunrespons.
TLR5 er til stede i immunsystemceller, så som lymfocytter, nøytrofiler, monocytter, makrofager, dendritiske celler, epitelceller og lymfatiske knuter. I tarmen regulerer TLR5 sammensetningen av mikrobiota.
Gram-negative bakterier bruker typisk Type-III sekretorisk system for å translokalisere svøpet til vertscellens cytoplasma, som utløser en serie intracellulære hendelser. Dermed gjenkjennes flagglinjen i det intracellulære miljøet av proteiner fra NAIP -familien (en hemmer av apoptose/NLR -familie).
Deretter samhandler Flagelina-NAIP5/6-komplekset med NOD-typen mottaker, som genererer vertens svar på infeksjon og skade.
Flagelina og planter
Planter gjenkjenner dette proteinet forresten Sensing 2 av Scourge (FLS2). Det siste er en kinase -reseptor i leucin repetisjoner og er en motstykke fra TLR5. FLS ”samhandler med den N-terminale regionen i LA Flagelina.
Union of Flagelin til FLS2 produserer fosforylering av kartkinaseveien, som kulminerer med syntesen av proteiner som formidler beskyttelse mot soppinfeksjon og bakterier.
I noen Solanáceas -planter kan svøpet også bli med i FLS3 -reseptoren. På denne måten er disse beskyttet mot patogener som unngår forsvaret mediert av FLS2.
Flagelina som adjuvans
Et adjuvans er et materiale som øker celle- eller humoral responsen på et antigen. Fordi mange vaksiner gir en dårlig immunrespons, er det nødvendig å ha gode adjuvanser.
Kan tjene deg: heterotrof bakterier: egenskaper og eksempler på arterTallrike studier demonstrerte effektiviteten av svøpe som adjuvans. Disse undersøkelsene besto av å bruke rekombinant flagelin i vaksiner, evaluert av dyremodeller. Imidlertid blir dette proteinet fortsatt overvunnet etter fase I av kliniske studier.
Blant de rekombinante flagelinasene er: Flagelina-epitop 1 av hematoglutinin av influensavirus; Flagelina-epitop Schistosoma Mansoni; Flageline-stabil varme til varme OG. coli; Flagelina -Proteína 1 av overflaten av Plasmodium; og Flagelina-protein fra Nile-virusinnpakningen, blant andre rekombinanter.
Det er noen fordeler med å bruke flagelin som adjuvans i vaksiner for menneskelig bruk. Disse fordelene er som følger:
1) Det er effektivt i veldig lave doser.
2) De stimulerer ikke IgE -svaret.
3) Du kan sette inn sekvensen til en annen adjuvans, AG, i svøpe -sekvensen uten å påvirke flaggelinjesignalet via TLR5.
Andre bruksområder av flagelin
Fordi flagelingenene viser bred variasjon, kan brukes til å utføre spesifikke deteksjoner, eller oppnå identifisering av arter eller stammer.
For eksempel har kombinasjonen av PCR/RFLP blitt brukt til å studere distribusjonen og polymorfismen av flagglinjegener i isolert OG. coli fra Nord -Amerika.
Referanser
- Hajam, i. TIL., Dar, s. TIL., Shahnawaz, i., Jaume, J. C., Les, J. H. 2017. Bakteriell flallin - et kraftimmunmodulerende middel. Eksperimentell og molekylær medisin, 49, E373.
- Kawamura-Sato, k., Inuma, og., Hasegawa, t., Horii, t., Yamashino, t., OHTA, m. 2000. Effekt av subinhibitorry konsentrasjoner av makrolider på ekspresjon av flamellin i Pseudomonas aeruginosa og Proteus mirabilis. Antimikrobielle midler og cellegift, 44: 2869-2872.
- Mizel, s. B., Bates, J. T. 2010. Flallin som en attjuvant: cellulære mekanismer og potensial. Journal of Immunology, 185, 5677-5682.
- Prescott, l. M., Harley, J. P., Klain, s. D. 2002. Mikrobiologi. Mc Graw-Hill, New York.
- Schaechter, m. 2009. Skrivebordet enyclopedia of Microbiology. Academic Press, San Diego.
- Winstanley, ca., Morgan, a. W. 1997. Bakteriell flamellinggen som biomarkør for deteksjon, populasjonsgenetikk og epidemiologisk analyse. Mikrobiologi, 143, 3071-3084.