

Aerobia glykolyse Hva er reaksjoner, glykolytiske mellommenn
- 2663
- 515
- Magnus Sander Berntsen
De Aerob eller aerob glykolyse Det er definert som bruk av glukoseoverskudd som ikke behandles ved oksidativ fosforylering mot dannelse av "fermentative" produkter, selv under forhold med høye oksygenkonsentrasjoner og til tross for fallet i energiytelsen.
Det er ofte gitt i vev med høye proliferative hastigheter, hvis forbruk av glukose og oksygen er høy. Et eksempel på dette er krefttumorceller, noen parasittiske celler av pattedyrblod og til og med cellene i noen områder av pattedyrhjerner.
Energien som er ekstrahert av katabolismen av glukose er bevart i form av ATP og NADH, som brukes nedstrøms i forskjellige metabolske veier.
Under aerob glykolyse er pyruvat rettet mot Krebs -syklusen og elektrontransportørkjeden, men den blir også behandlet av Fermentative for regenerering av NAD+ uten ytterligere ATP -produksjon, som ender med laktatdannelse.
Aerob eller anaerob glykolyse forekommer hovedsakelig i cytosol, med unntak av organismer som Tripanosomatider, som har spesialiserte glykolytiske organeller kjent som glykosomer.
Glykolyse er en av de mest kjente metabolske traséene. Det ble fullstendig formulert på 1930 -tallet av Gustav Embden og Otto Meyerhof, som studerte ruten i skjelettmuskelceller. Aerob glykolyse er imidlertid kjent som Warburg -effekten siden 1924.
Aerobe glykolysereaksjoner
Aerob katabolisme av glukose skjer i ti katalyserte trinn. Mange forfattere vurderer at disse trinnene er delt inn i en energiinvesteringsfase, som tar sikte på å øke gratis energiinnholdet i mellommenn, og en annen erstatning og ATP -formet energiforsterkning.
Kan tjene deg: histokjemi: fundament, prosessering, fargingEnergiinvesteringsfase
1-fosforylering av glukose til glukose 6-fosfat katalysert av heksokinase (HK). I denne reaksjonen investeres den, for hvert glukosemolekyl, et ATP -molekyl, som fungerer som en fosfatgruppedonor. Glukose 6-fosfat (G6P) og ADP, og reaksjonen er irreversibel.
Enzymet krever dannelse av en komplett Mg-ATP2- for dens drift, så det fortjener magnesiumioner.
2-uomerisering av G6P til fruktose 6-fosfat (F6P). Det involverer ikke energiutgifter og er en reversibel reaksjon katalysert av fosfoglukoseisomerase (PGI).
3-f6p fosforylering til fruktose 1,6-biffosfat katalysert av fosfofratoquinase-1 (PFK-1) (PFK-1). Et ATP-molekyl brukes som en fosfatgruppedonor, og reaksjonsproduktene er F1,6-bp og ADP. Takket være verdien av ∆G er denne reaksjonen irreversibel (som reaksjon 1).
4-katalytikopur av F1.6-bp i dihydroksyacetonfosfat (DHAP), en ketose og glyseraldehyd 3-fosfat (GAP), en aldosa. Aldolase -enzym er ansvarlig for denne reversible aldoliske kondensasjonen.
5-Triosa-fosfatisomerase (TIM) er ansvarlig for interkonversjonen av triosas fosfat: DHAP og gapet, uten ekstra energibidrag.
Energigjenvinningsfase
1-Gapet oksideres av glyseraldehyd 3-fosfatdehydrogenase (GAPDH), som katalyserer overføringen av en gap fosfatgruppe for å danne 1,3-bifoskglyceratet. I denne reaksjonen reduseres to NAD+ molekyler med glukosemolekyl, og to uorganiske fosfatmolekyler brukes.
Hver NADH produserte passerer gjennom elektrontransportkjeden og 6 ATP -molekyler syntetiseres ved oksidativ fosforylering.
2-fosfoglyseratkinase (PGK) overfører en fosforylgruppe fra 1,3-bifosfoglyserat til ADP, og danner to ATP-molekyler og to av 3-fosfoglyserat (3pg) (3pg). Denne prosessen er kjent som fosforylering på underlagsnivå.
Kan tjene deg: Stabilisatorvalg: Hva er og eksemplerDe to ATP -molekylene som konsumeres i HK- og PFK -reaksjonene blir besvart av PGK i denne passasjen av ruten.
3-3pg omdannes til 2pg av mutasa fosfoglyserat (PGM), som katalyserer forskyvningen av fosforylgruppen mellom karbon 3 og 2 av glyseratet i to trinn og reversibelt. Magnesiumionet er også nødvendig av dette enzymet.
4-A dehydreringsreaksjon katalysert av enolasas konverterer 2pg til fosfoenolpyruvat (PEP) til en reaksjon som ikke krever energiinvestering, men genererer en forbindelse med større energipotensial for overføring av fosfatgruppen senere.
5-Finalt, kinase Pyruvate (PYK) katalyser. To ADP -molekyler brukes av glukosemolekyl og 2 ATP -molekyler genereres. Pyk bruker kalium- og magnesiumioner.
Dermed er den totale energiytelsen til glykolyse 2 ATP -molekyler for hvert glukosemolekyl som kommer inn i ruten. Under aerobe forhold innebærer fullstendig glukosedegradering å oppnå mellom 30 og 32 ATP -molekyler.
Destinasjon for glukolitiske mellommenn
Deretter blir glykolyse, pyruvatet utsatt for dekarboksylering, produserer CO2 og donerer acetylgruppen til acetylkoenzym A, som også er oksidert til CO2 i Krebs -syklusen.
Elektroner som frigjøres under denne oksidasjonen blir transportert til oksygen gjennom reaksjonene fra den mitokondrielle respirasjonskjeden, som til slutt fremmer syntesen av ATP i denne organellen.
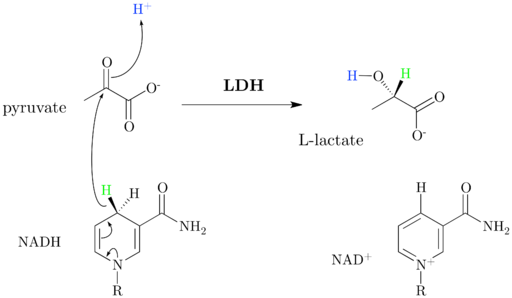
Det kan tjene deg: Flora og Fauna de Salta: Mer representative arterUnder aerob glykolyse behandles overflødig pyruvat som produseres av enzymet laktatdehydrogenase, som danner laktat og regenererer en del av NAD+ konsumert trinn opp i glykolyse, men uten dannelse av nye ATP -molekyler.
Laktatdehydrogenasemekanisme (Kilde: Jazzlw [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) I tillegg kan pyruvat brukes i anabole prosesser som fører til dannelse av alaninaminosyre, for eksempel, eller også kan fungere som et skjelett for fettsyresyntese.
I likhet med Pyruvate, det endelige produktet av glykolyse, oppfyller mange av reaksjonsformidlere andre funksjoner på kataboliske eller anabolske ruter som er viktige for cellen.
Slik er tilfellet med glukose 6-fosfat og ruten til pentosefosfat, der mellommennene til ribosøs som er til stede i nukleinsyrer oppnås.
Referanser
- Akram, m. (2013). Mini-anmeldelse om glykolyse og kreft. J. CANC. Utdanning., 28, 454-457.
- Esen, e., & Lang, f. (2014). Aerob glykolyse i osteoblaster. Curr Osteoporos Rep, 12, 433-438.
- « Efebophilia hva er, symptomer, årsaker, konsekvenser, behandling
- Isomerase hva er, funksjoner, nomenklatur, typer »