

Anaerob glykolyse Hva er reaksjoner, fermentative stier
- 2119
- 214
- Marius Aasen
Hva er anaerob glykolyse?
De Anaerob glykolyse o Anaerob er en katabolsk rute som brukes av mange typer celler for glukosedbrytning i fravær av oksygen. Det vil si at glukose ikke er fullstendig oksidert til karbondioksid og vann, som tilfellet er med aerob glykolyse, men fermentative produkter genereres.
Det kalles anaerob glykolyse, siden det finner sted uten tilstedeværelse av oksygen, som i andre tilfeller fungerer som den endelige elektronakseptoren i mitokondriene transportørkjeden, der store mengder energi produseres fra behandlingen av glykolytiske produkter.
Avhengig av organismen, vil en anaerobiosetilstand eller fravær av oksygen resultere i produksjon av melkesyre (muskelceller, for eksempel) eller etanol (gjær), fra pyruvatet generert ved katabolisme av glukose.
Som et resultat faller energiytelsen dramatisk, siden bare to mol ATP produseres for hver mol glukose som behandles, sammenlignet med de 8 molene som kan oppnås under aerob glykolyse (bare i glykolytisk fase).
Forskjellen i antall ATP -molekyler har å gjøre med reoksidasjonen av NADH, som ikke genererer ytterligere ATP, i motsetning til hva som skjer i aerob glykolyse, som for hver NADH oppnådde 3 ATP -molekyler.
Reaksjoner
Anaerob glykolyse kommer overhodet ikke fra aerob glykolyse, siden begrepet "anaerob" snarere refererer til hva som skjer etter den glykolytiske ruten, det vil si til reaksjonsprodukter og mellomleddsmål og formidlere.
Kan tjene deg: luftrøretI reaksjonene fra anaerob glykolyse deltar således ti forskjellige enzymer, nemlig:
1-heksokinase (HK): Bruk et ATP-molekyl for hvert glukosemolekyl. Produserer glukose 6-fosfat (G6P) og ADP. Reaksjonen er irreversibel og fortjener magnesiumioner.
2-Fosfoglucoso isomerasa (PGI): Isomeriza G6P A fruktose 6-fosfat (F6P).
3-fosfofrucerachinase (PFK): Fosphoryila F6p A fruktose 1,6-bifosfat (F1,6-bp) ved bruk av et ATP-molekyl for hver F6P, denne reaksjonen er også irreversibel.
4-Aldolase: Escind F1.6-bp-molekylet og produserer glyseraldehyd 3-fosfat (GAP) og dihydroksyaceton fosfat (DHAP).
5-fosfatisomerase (TIM): Deltar i DHAP og GAP-interkonvertering.
6-glyseraldehyd 3-fosfatdehydrogenase (GAPDH): Bruk to NAD-molekyler+ og 2 uorganiske fosfatmolekyler (PI) for fosforylering av gapet, gir 1,3-bifosfoglyserat (1,3 bpg) og 2 NADH.
7-Fosfoglicerato Quinasa (PGK): produserer to ATP-molekyler på grunn av fosforylering ved underlagsnivået til to ADP-molekyler. Vekt med fosfatgruppegiver hvert 1,3 bpg molekyl. Produserer 2 3-fosfoglyserat (3pg) molekyler.
8-Fosfoglicerato mutasa (PGM): Oorganiser 3pg-molekylet for å forårsake en mellomledd med større energi, 2pg.
199.
10-pyruvat kinase (PYK): Fosfoenolpyruvat brukes av dette enzymet for å danne pyruvat. Reaksjonen innebærer overføring av fosfatgruppen i posisjon 2 i fosfoenolpiruvat til et ADP -molekyl. 2 pyruvater og 2 ATP produseres for hver glukose.
Fermentative ruter
Fermentering er betegnelsen som brukes for å indikere at glukose eller andre næringsstoffer blir nedbrutt i fravær av oksygen, for å oppnå energi.
I mangel av oksygen har ikke elektrontransportørkjeden en endelig akseptor og forekommer derfor ikke oksidativ fosforylering som betaler store mengder energi i form av ATP. NADH er ikke ReoxyD med mitokondriell rute, men med alternative ruter, som ikke produserer ATP.
Det kan tjene deg: Pyruvate kinase: struktur, funksjon, regulering, hemmingUten nok NAD+ Den glykolytiske banen.
Noen celler har alternative mekanismer for å takle perioder med anaerobiose, og generelt innebærer disse mekanismene en eller annen type gjæring. Andre celler, tvert imot, avhenger nesten utelukkende av fermentative prosesser for livsopphold.
Produktene fra de gjærende veiene til mange organismer er økonomisk relevante for mennesket; Eksempler er produksjon av etanol av noen gjær i anaerobiose og dannelse av melkesyre ved laktbakterier brukt til yoghurtproduksjon.
Melkesyreproduksjon
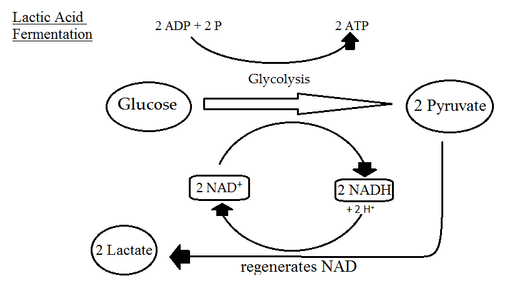
Mange typer celler i fravær av oksygen produserer melkesyre takket være reaksjonen katalysert av dehydrogenase laktatkompleks, som bruker pyruvatkarbonene og NADH produsert i GAPDH -reaksjonen.
Laktisk gjæring (kilde: Sjantoni [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) Etanolproduksjon
Pyruvatet omdannes til acetaldehyd og CO2 ved pyruvatdekarboksylase. Acetaldehyd blir deretter brukt av alkoholdehydrogenase, noe som reduserer det ved å produsere etanol og regenerere et NAD -molekyl+ for hvert pyruvatmolekyl som kommer inn på denne måten.
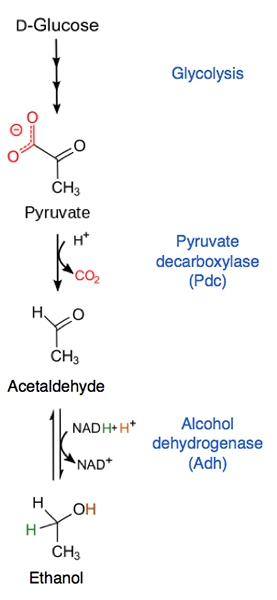 Alkoholholdig gjæring (kilde: AROBSON1 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Alkoholholdig gjæring (kilde: AROBSON1 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Aerob gjæring
Anaerob glykolyse har som hovedkarakteristisk det faktum at sluttprodukter ikke tilsvarer CO2 og vann, som i tilfelle av aerob glykolyse. I sin plass genereres typiske gjæringens reaksjoner.
Noen forfattere har beskrevet en prosess med "aerob gjæring" eller aerob glukly glykolyse for visse organismer, blant dem noen parasitter i Trypanosomatidae -familien og mange kreftcreftceller skiller seg ut.
Det kan tjene deg: termoreceptorer: hos mennesker, i dyr, i planterI disse organismer har det blitt vist at selv i nærvær av oksygen er produktene fra glykolytisk vei mulig for karbonatomer.
Selv om "aerob gjæring" av glukose ikke innebærer det totale fraværet av luftveisaktivitet, siden det ikke er en prosess av alle eller ingenting. Imidlertid indikerer bibliografien utskillelse av produkter som pyruvat, laktat, succinat, ond og andre organiske syrer.
Glykolyse og kreft
Mange kreftceller viser en økning i glukose og glykolytisk strømning.
Svulster hos kreftpasienter vokser raskt, så blodkar er i hypoksi. Dermed avhenger energitilskuddet til disse cellene hovedsakelig av anaerob glykolyse.
Imidlertid hjelper dette fenomenet av en hypoksiinduserbar transkripsjonsfaktor (HIF), noe som øker ekspresjonen av glykolytiske enzymer og glukosetransportører i membranen gjennom komplekse mekanismer.
Referanser
- Cazzulo, J. J. (1992). Aerob gjæring av glukose av trypanosomatider. FASB Journal, 6, 3153-3161.
- Jones, w., & Bianchi, K. (2015). Aerob glykolyse: utover spredning. Grenser i immunologi, 6, 1-5.