

Mikrofilamenter Karakteristikker, struktur, funksjoner, patologi
- 2955
- 721
- Mathias Aas
De mikrofilamenter o Aktinfilamenter, er en av de tre grunnleggende komponentene i eukaryotisk cellcytoskelet.
I eukaryoter er genene som koder for aktinmikrofilamenter sterkt bevart i alle organismer, så de blir ofte brukt som molekylære markører for forskjellige studier.
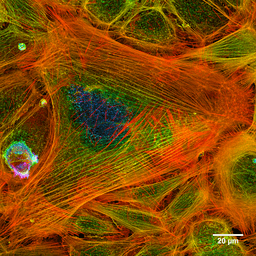
Fotografi av aktinfilamentene til en farget celle (kilde: Howard Vindin [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Mikrofilamenter er fordelt over cytosol, men de er spesielt rikelig i det underliggende området til plasmamembranen, hvor de danner et komplekst nettverk og er assosiert med andre spesielle proteiner for å danne cytoskjelettonet.
Mikrofilamentnettverk i cytoplasma av pattedyrceller er kodet av to av de seks genene som er beskrevet for aktin, som er involvert i dynamikken i mikrofilamenter og som til og med er veldig viktige under differensieringen av stamcellene.
Mange forfattere er enige om at mikrofilamenter er de mest forskjellige, allsidige og viktige proteiner i cytoskjelettet til de fleste eukaryote celler, og det er viktig å huske at disse ikke finnes i prokaryote mikroorganismer.
I denne typen celler er det imidlertid homologe filamenter med mikrofilamenter, men som dannes av et annet protein: MREB -protein.
For tiden antas det at genet som koder for dette proteinet er et mulig forfedres gen for eukaryotisk aktin. Imidlertid er aminosyresekvenshomologien som danner MREB -protein bare 15% med hensyn til aktinsekvensen.
Som en grunnleggende del av cytoskjelettet, kan enhver fenotypisk defekt både i mikrotubuli og i mellomliggende filamenter og aktinmikrofilamenter (cytoskjelett) forårsake forskjellige celle- og systemiske patologier.
[TOC]
Egenskaper og struktur
Mikrofilamenter består av proteinmonomerer av aktinfamilien, som er ekstremt rikelig med kontraktile proteiner i eukaryote celler, siden de også deltar i muskelsammentrekning.
Disse filamentene har en diameter mellom 5 og 7 nm, så de er også kjent som tynne filamenter og er sammensatt av to former for aktin: den kuleformen (aktin G) og den filamentøse formen (aktin F).
Proteiner som deltar i cytoskjelettet er kjent som γ og ß -aktus, mens de som deltar i sammentrekning vanligvis er aktiner α.
Kan tjene deg: cytoplasma: funksjoner, deler og egenskaperAndelen kuleaktin og filamentøs aktin i cytosol avhenger av cellulære behov, siden mikrofilamenter er veldig varierende og allsidige strukturer, som kontinuerlig er vekst og forkortelse på grunn av polymerisasjon og depolymetisering.
Aktin G er et lite kuleformet protein, bestående av nesten 400 aminosyrer og rundt 43 kDa molekylvekt.
Aktin G -monomerer som utgjør mikrofilamentene er bestilt i form av en spiralformet streng, siden hver enkelt lider av en torsjon når de er assosiert med følgende.
Aktin G er assosiert med et Ca2+ -molekyl og en annen ATP, som stabiliserer dens kuleform; Mens aktin F oppnås etter hydrolyse av terminalfosfatet til ATP -molekylet i aktin G, noe som bidrar til polymerisasjon.
Organisasjon
Aktinfilamenter kan organiseres i form av "bjelker" eller "nettverk" som har forskjellige funksjoner i cellene. Strålene danner parallelle strukturer knyttet sammen med ganske stive kryssbroer.
Nettverk er derimot mer komfortable strukturer, for eksempel tre -dimensjonale masker med egenskapene til semi -solide geler.
Det er mange proteiner som er assosiert med aktin- eller mikrofilamentfilamenter, og som er kjent som ABP (fra engelsk Aktinbindende proteiner), som har spesifikke nettsteder for dette.
Mange av disse proteinene lar mikrofilamenter samhandle med de to andre komponentene i cytoskjelettet: mikrotubuli og mellomliggende filamenter, så vel som med de andre komponentene i den indre ansiktet til plasmamembranen.
Blant andre proteiner som interagerer med mikrofilamenter, er kjernefysiske ark og spektrin (i røde blodlegemer).
Hvordan dannes aktinfilamenter?
Siden kuleformede aktinmonomerer alltid blir med på samme måte, orientert i samme retning, har mikrofilamenter en definert polaritet, med to ytterpunkter: en "mer" og en "mindre".
Polariteten til disse filamentene er veldig viktig, siden de vokser betydelig raskere på grunn av deres positive slutt, der de nye aktinene er tilsatt.
 Grafisk representasjon av aktinmikrofilamenter (kilde: Derivatarbeid: retama (snakk) tynn_filament_formasjon.SVG: Mikael Häggström [Public Domain] via Wikimedia Commons)
Grafisk representasjon av aktinmikrofilamenter (kilde: Derivatarbeid: retama (snakk) tynn_filament_formasjon.SVG: Mikael Häggström [Public Domain] via Wikimedia Commons) Det første som foregår under polymerisering av aktinfilamenter er en prosess kjent som "kjernefysning", som består av assosiasjonen til tre proteinmonomerer.
Kan tjene deg: plasomolyseTil denne trimmeren blir nye monomerer lagt til i begge ender, slik at glødetråden vokser. Aktin G-monomerer er i stand til å hydrolyz ATP med hver forening, noe som har implikasjoner i hastigheten på polymerisasjon, siden aktin-ATP-porsjoner dissosierer mer vanskeligheter enn aktin-ADP.
ATP er ikke nødvendig for polymerisasjon, og den konkrete funksjonen til dens hydrolyse er ennå ikke avklart.
Noen forfattere vurderer det, siden aktinpolymerisasjonshendelser er raske.
Regulering
Både polymerisering av aktinfilamenter og dens depolimerisering er prosesser som er sterkt regulert av en serie spesifikke proteiner, som er ansvarlige for ombygging av filamentene.
Eksempel på proteinene som regulerer depolymerer. Et annet protein, profylain, har en motsatt funksjon, siden det stimulerer assosiasjonen av monomerer (ved å stimulere ADPs utveksling for ATP).
Funksjoner
Mikrofilamenter interagerer med myosinfilamenter som er assosiert med transmembranproteiner som har et domene i cytosol og en annen i utlandet celle ute, så de deltar i cellemobilitetsprosesser.
Disse mikrofilamentene assosiert med plasmamembranen medierer forskjellige cellulære responser til forskjellige typer stimuli. For eksempel er celleadhesjon av epitelvev rettet av transmembranproteiner kjent som cadherinas, som samhandler med mikrofilamenter for å rekruttere responsfaktorer.
Aktinfilamenter interagerer med mellomliggende filamenter for å få ekstracellulære stimuli overføring til viktige steder som ribosomer og kromosomer inne i kjernen.
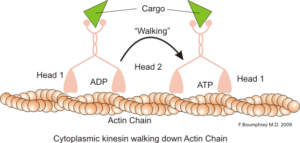 Representasjon av den intracellulære motoriske funksjonen til aktinmikrofilamenter (kilde: Boumphreyfr [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons)
Representasjon av den intracellulære motoriske funksjonen til aktinmikrofilamenter (kilde: Boumphreyfr [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) En klassisk og veldig studert funksjon av mikrofilamenter er dens evne til å danne "broer", "skinner" eller "motorveier" for bevegelse av myosinmotorprotein I, som er i stand til å laste transportvesikler fra organellene til membranplasmaet på sekretorien stier.
Mikrofilamenter interagerer også med myosin II for å etablere den kontraktile ringen som dannes under cytokinesis, nettopp i løpet av det siste trinnet av celledelingen der cytosol er atskilt fra stam- og dattercellene.
Kan tjene deg: polymorfonukleære leukocytterGenerelt modulerer aktin -formede mikrofilamenter fordelingen av noen organeller som Golgi -komplekset, endoplasmatisk retikulum og mitokondrier. I tillegg deltar de også i den romlige plasseringen av RNMS slik at de blir lest av ribosomene.
Hele mobiltelefonsettet med mikrofilamenter, spesielt de som er nært beslektet med plasmamembranen, deltar i dannelsen av bølgende cellemembraner av celler som har en konstant aktiv bevegelse.
De deltar også i dannelsen av mikrovinger og andre vanlige fremspring på overflaten av mange celler.
Eksempel på funksjoner i leveren
Mikrofilamenter deltar i gallesekresjonsprosessen i hepatocytter (leverceller) og også i peristaltiske bevegelser (koordinert sammentrekning) av leverkanalikuli.
De bidrar til differensiering av plasmamembrandomener takket være deres tilknytning til forskjellige cytosoliske elementer og kontrollen de utøver på topografien til disse intracellulære elementene.
Relaterte patologier
Det er få sykdommer assosiert med primære defekter i strukturen eller med proteiner og regulatoriske enzymer i syntesen av mikrofilamenter, selv om disse er direkte involvert i stort antall funksjoner.
Den lave indeksen for sykdommer og misdannelser i den primære strukturen til mikrofilamenter skyldes det faktum at det generelt er flere kodingsgener av både aktin og dets regulatoriske proteiner, et fenomen kjent som "genetisk redundans".
En av de mest studerte patologiene er forklingning av oocytter på deres cytoskjelett, hvor et avbrudd observeres i det kortikale mikrofilamentnettverket, så vel som en depolimerisering og en uorganisering av mikrotubulene i den mitotiske spindelen.
Generelt sett forårsaker denne forklingingen kromosomisk spredning, siden den fører til en uenighet i komprimeringen av hele kromatinet.
Cellene som har en større organisering og andel mikrofilamenter i cytoskjelettet er stripete muskelceller, derfor er de fleste patologier assosiert med en funksjonsfeil i det kontraktile apparatet.
Mangelfulle eller atypiske mikrofilamenter har også vært assosiert med beinsykdom kjent som Pagets sykdom.
Referanser
- Aguilar-Cuenca, R., Llorente-González, ca., Vicente, ca., & Vicente-Manzanares, M. (2017). Mikrofilament-koordinert vedheftingsdynamikk driver migrasjon av enkeltceller og former WHOE-vev. F1000Resarch, 6.
- To midler, C. G., Chhabra, d., Kekic, m., Finger, i. V., Tsubakihara, m., Berry, d. TIL., & Nosworthy, n. J. (2003). Bindende proteiner aktin: regulering av cytoskeletale mikrofilamenter. Fysiologiske anmeldelser, 83(2), 433-473.
- Guo, h., Fauci, l., Shelley, m., & Kanso, og. (2018). Bistabilitet i synkroniseringen av aktiverte mikrofilamenter. Journal of Fluid Mechanics, 836, 304-323.
- Lansering., Langer, r., & Vacanti, J. P. (Eds.). (2011). Prinsipper for vevteknikk. Akademisk presse.
- Robbins, J. (2017). Sykdommer i cytoskjelettet: Deminopatiene. Hos kardioskeletale myopatier hos barn og dere voksne (PP. 173-192). Akademisk presse.
- « Grizzly bjørnegenskaper, habitat, mat, oppførsel
- Cistus clusii -egenskaper, taksonomi, habitat, distribusjon, bruk »