

Fluidmosaikkmodell
- 2559
- 326
- Prof. Joakim Johansen
Vi forklarer hva væskemosaikkmodellen, strukturen til cellemembranen og dens funksjoner er.
Fluid Mosaic Model Structure Hva er fluidmosaikkmodellen?
Han Fluidmosaikkmodell Den sier at celle- eller biomembranmembraner er dynamiske strukturer som har flyt av sine forskjellige molekylære komponenter, som kan bevege seg sideveis. Det vil si at disse komponentene er i bevegelse og ikke -statisk, som tidligere antatt.
Denne modellen ble hevet av S. Jonathan Singer og Garth. L. Nicolson i 1972 og er i dag allment akseptert av det vitenskapelige samfunnet. Alle celler er inneholdt av en cellemembran med særegenheter i dens konstitusjon og funksjon.
Denne membranen definerer cellens grenser, og tillater eksistensen av forskjeller mellom cytosol (eller cellulært interiør) og det ytre miljøet. Den regulerer også utveksling av stoffer mellom cellen og utsiden.
I eukaryote celler definerer indre membraner også rom og organeller med forskjellige funksjoner, for eksempel mitokondrier, kloroplaster, kjernefysisk konvolutt, endoplasmatisk retikulum, Golgis apparat, blant andre.
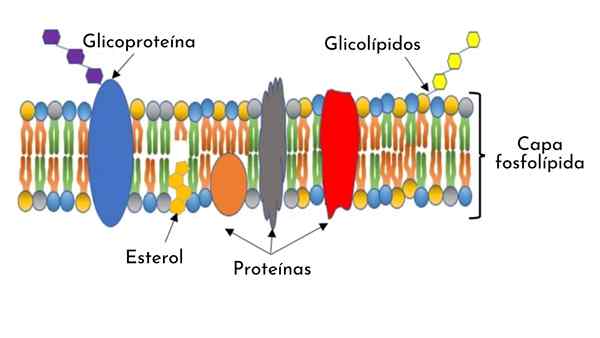
Cellemembranstruktur
Generaliteter
- Cellemembranen består av en vanntett struktur til molekyler og vannoppløselige ioner mellom 7 og 9 nanometer tykke. Det observeres i elektroniske mikrofotografier som en dobbel kontinuerlig og tynn linje som omgir cellens cytoplasma.
- Den er sammensatt av et fosfolipid -dobbeltlag, med proteiner innebygd langs strukturen og arrangert på overflaten.
- Den inneholder karbohydratmolekyler på begge overflater (indre og eksterne), og når det.
Fosfolipid -dobbeltlag
Fosfolipider er amfipatiske molekyler som har en hydropofisk ende -vannfin -og en annen hydrofob -som frastøter vann-.
Fosfolipid -dobbeltlaget som danner cellemembranen har hydrofobe (apolare) kjeder arrangert inne i membranen, og hydrofile (polare) ytterpunkter lokalisert mot det ytre medium.
Det kan tjene deg: Flora og fauna fra USADermed blir hodene til fosfatgrupper av fosfolipider utsatt på den ytre overflaten av membranen.
Husk at både den ytre og den interne eller cytosolen er vandig. Dette påvirker arrangementet av det doble fosfolipidlaget, med dets polare deler som samhandler med vann og dets hydrofobe deler som danner den indre membranmatrisen.
Kolesterol
I membranen til Eukaryot dyreceller er det kolesterolmolekyler satt inn i de hydrofobe halene til fosfolipider.
Disse molekylene finnes ikke i membranene til prokaryote celler, noen protister, planter og sopp.
Omfattende membran- eller transmembranproteiner
Inne i fosfolipid Bilay.
Disse samhandler ikke kovalent gjennom sine hydrofobe deler, med lipid bilay.
Membranproteinkonfigurasjon
De kan presentere en enkel konfigurasjon i form av en baston, med et alfa hydrofob og innebygd alfa -propell inne i membranalt interiør, og med utvidede hydrofile deler til sidene.
De kan også presentere en større konfigurasjon av kuletype, og med en kompleks tertiær eller kvartær struktur.
Sistnevnte krysser ofte cellemembranen flere ganger med sine alfa -propellsegmenter gjentatt og anordnet i sikksakk gjennom lipid -dobbeltlaget.
Porer i membranene
Noen av disse kuleproteinene har hydrofile interiørdeler, og danner kanaler eller porer som utvekslingen av polare stoffer oppstår fra cellen utenfor til cytosol, og omvendt.
Perifere proteiner
På overflaten av det cytoplasmatiske ansiktet til cellemembranen er det perifere membranproteiner, knyttet til de enestående delene av noen integrerte proteiner.
Det kan tjene deg: Aptenia cordifolia: Kjennetegn, habitat, egenskaper, dyrkingDisse proteinene trenger ikke gjennom den hydrofobe kjernen til lipid -dobbeltlaget.
Karbohydratdekke
Det er karbohydratmolekyler på begge membranoverflater.
Spesielt har den ytre overflaten av membranen en overflod av glukolipider. Korte karbohydratkjeder blir også observert eksponert og kovalent festet til de enestående proteindelene, kalt glykoproteiner.
Cellemembranfluiditet
Andel mettede fettsyrer vs. umettet
Fluiditeten til membranen avhenger hovedsakelig av andelen mellom fosfolipider av mettede og ikke -mettede fettsyrer som er til stede.
Denne membranalfluiditeten avtar når andelen fosfolipider av mettede fettsyrekjeder øker, med hensyn til ikke -mettet.
Dette er fordi samhold er større mellom lange og enkle kjeder av mettede fettsyrer, med hensyn til samhold mellom de korte og umettede kjederene av umettede fettsyrer.
En større samhold blant molekylære komponenter, lavere flyt vil presentere membranen.
Kolesterol
Kolesterolmolekyler interagerer gjennom sine stive ringer med lipidhydrokarbonkjeder, noe som øker membranens stivhet og reduserer permeabiliteten til det samme.
I membranene til flertallet av eukaryote celler, der det er en relativ høy kolesterolkonsentrasjon, forhindrer dette kullsyreholdige kjeder i å slå sammen lave temperaturer. Dette forventes å fryse membranen ved lave temperaturer.
Særegenheter
De forskjellige typene cellemembraner presenterer særegenheter i deres mengde og type protein og karbohydrater, så vel som i mangfoldet av eksisterende lipider.
Disse særegenhetene er assosiert med spesifikke cellulære funksjoner.
Det er ikke bare konstitutive forskjeller mellom eukaryot og prokaryote cellemembraner og blant organellene, men også mellom regioner med samme membran.
Kan tjene deg: SSB -proteiner: Kjennetegn, struktur og funksjonerCellemembranfunksjoner
- Cellemembranen avgrenser cellen og lar den opprettholde en stabil tilstand i cytosol, forskjellig fra det ytre miljøet.
- Gjennom aktiv og passiv regulering av passering av stoffer (vann, ioner og metabolitter) gjennom seg selv, opprettholder den det nødvendige elektrokjemiske potensialet for cellulær drift.
- Den lar cellen svare på eksterne mediumsignaler gjennom kjemiske membranreseptorer og gir forankringssteder for cytoskjelettfilamenter.
- Når det gjelder eukaryote celler, deltar den også i etablering av interne rom og organeller med spesifikke metabolske funksjoner.
Proteinfunksjon i membranen
Det er forskjellige membranproteiner med spesifikke funksjoner, blant dem vi kan nevne:
- Enzymer som katalyserer (akselererer) kjemiske reaksjoner.
- Membranreseptorer som deltar i anerkjennelse og union til signalmolekyler (for eksempel hormoner).
- Stofftransportproteiner gjennom membranen (til cytosol og fra dette til celleutvendig). Disse opprettholder en elektrokjemisk gradient takk til ionetransport.
Eksternt karbohydratdekkefunksjon
Karbohydrater eller glukolipider deltar i vedheft av celler med hverandre og i prosessen med gjenkjennelse og interaksjon av cellemembran med molekyler som antistoffer, hormoner og virus.
Referanser
- Bagsover, s. R., Hyams, J. S., Shephard, e. TIL., Hvit, h. TIL. og Wiedemann, C. G. (2003). Cellebiologi, til kort kurs. Andre utgave. Wiley-Liss.
- Engelman, d. (2005). Er mer mosaikk er flytende membraner. Nature 438 (7068).
- Raven, J. (2002). Biologi. Sjette utgave. Mgh.