

Peyer -plater egenskaper, funksjoner, histologi
- 3122
- 201
- Oliver Christiansen
De Peyer -plater De er anatomiske regioner som ligger under slimhinnen i mage -tarmkanalen, spesielt i tynntarmens lille ark. De er aggregeringssteder for et stort antall lymfocytter og andre tilbehørsceller, slik at de representerer en del av immunforsvaret i slimhinnene.
I likhet med mandlene i svelget og at lymfoide folliklene i substankenes submukos.
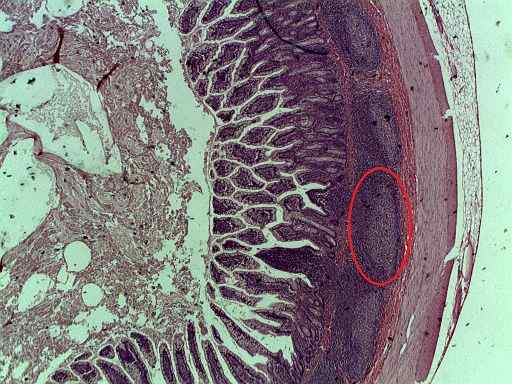
 Fotografi av Peyers plater i et tverrsnitt av tynntarmen (Kilde: Plainpaper [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons)
Fotografi av Peyers plater i et tverrsnitt av tynntarmen (Kilde: Plainpaper [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) Det er viktig å huske at immunresponsen (kroppens forsvarssystem mot "inntrengerne" eksterne midler) er mediert av forskjellige typer celler, og er lymfocytter av det viktigste, fordi de takket være dens evne til å gjenkjenne antigener, er de bestilt å utløse de spesifikke immunresponsene.
Peyers plater ble beskrevet i 1645 som "lymfoide follikler" av den italienske Marco Aurelio Severino, men det var først i 1677 at "Peyer Plates" ble myntet til ære for den sveitsiske patologen Johann Conrad Peyer, som beskrev en beskrivelse detaljert detaljert detaljert detaljert.
Funksjonen ble imidlertid bestemt mange år senere da Kenzaburo Kumagai i 1922 la merke til "absorpsjon" -kapasiteten til patogene og fremmede celler fra epitel til epitelens "kuppel" av peyerplatene.
[TOC]
Kjennetegn
Peyers plater tilhører det som er kjent som "lymfoidvevet assosiert med tarmen" eller Galt (fra engelsk Gut-TILSsociated LYmfoid TProblem "), som er sammensatt av lymfoide follikler fordelt langs mage -tarmrøret.
Dette lymfoide vevet assosiert med tarmen representerer et av de største lymfoide organene i kroppen, siden det inneholder nesten 70% av immunocytt eller "immunocytt" -celler.
En lymfoid follikkel er et aggregat eller sett med lymfoide celler som ikke har en klar struktur eller en bestemt organisasjon.
General.
I den menneskelige tynntarmen har Peyers plater en "oval" form og er uregelmessig distribuert. Cornes, i 1965, bestemte at antall plater under menneskelig utvikling har en topp mellom 15 og 25 år og deretter avtar med alderen.
Andre forskere har forsikret at området okkupert av Peyers plater i ileum har et maksimalt punkt i løpet av det tredje tiåret av livet, og at den største andelen av disse er konsentrert i løpet av de siste 25 cm av ileum.
Kan tjene deg: kort radial extensor av carpo: anatomi, funksjoner, lateral epikondylittSom mange andre vev i menneskekroppen, avhenger organogenesen av Peyers plater i stor grad av deltakelse av spesifikke cytokiner som formidler differensiering og arrangement av disse anatomiske regionene.
Funksjoner
Hovedfunksjonen til Peyer -platene som en del av immunforsvaret i tarmslimhinnene er å beskytte "skallet" mot tarmen i invasjonen ved potensielt patogene mikroorganismer.
Noen av cellefolliklerceller som er til stede i denne "regionen" i tarmen er ansvarlige for å skille mellom patogene mikroorganismer og "spisestue" (som tilhører den native mikroflora), siden disse folliklene samhandler direkte med tarmpitel.
I induksjon av "oral" immuntoleranse
Peyerplater deltar i "absorpsjonen" av fremmed- eller patogene celler, men det er vist at celler som tilhører nevnte region også er i stand til å skille mellom visse antigener og mellom ikke -patogene bakterier assosiert med tarmkanalen.
Denne ikke -patogene gjenkjennelsesprosessen er kjent som "oral toleranse", og den er en aktiv prosess som fører til dannelse av spesifikke T -lymfocytter som er i stand til å unngå utløseren av en unødvendig immunrespons.
Oral toleranse er også definert som den antigenspesifikke eliminering av humorale og cellulære immunresponser på antigener som når kroppen ved oral rute, og er spesielt nyttig for beskyttelse av tarmslimhinnen i møte med ugunstige inflammatoriske immunresponser.
Histologi
Peyers plater er en del av det lille arket i tynntarmen. Det eget arket er sammensatt av det slappe bindevevet som samtidig er en del av det som kalles "kjernen" av tarmvilli.
I det eget arket er det forskjellige typer plasmaceller, lymfocytter, leukocytter, fibroblaster, byggceller og andre, og Peyers plater er den delen av sine egne laminaer der permanente sett med knuter eller lymfolfollikler er funnet.
- Struktur
Peyers plater skilles arkitektonisk inn i tre hoveddomener kjent som:
1- Follikulære området
2- Det grensede området og
3- Epitel assosiert med lymfoide follikler.
Follikulært og interfolært område
Denne regionen består av nodulene eller lymfoide folliklene på peyerplatene som består av B -celler (lymfocytter B) omgitt av en mindre kompakt del (LAXA) av T -celler (T -lymfocytter) og mange follikulære dendritiske celler eller “antigenpresentant) celler ”(APC, engelsk TILntigen PHarme CElls).
Den delen der lymfocytter eller replikative B -celler, dendritiske celler og andre celler, makrofager kalles "Germinal Center". Hver lymfoide follikkel er på sin side omgitt av det som er kjent som "korona" eller "subepitelkuppel" ".
Kan tjene deg: urinsystemDen subepitelkuppelen har også en blanding av lymfoide celler (B- og T -lymfocytter), follikulære og makrofag -dendritiske celler, og det er dette det grensede området representerer.
Det er vist at i lymfoide follikler av voksne mus er andelen B -celler i det indre området av disse strukturene omtrent 50 eller 70%, mens T -celler bare representerer 10 til 30%.
Noen undersøkelser antyder også tilstedeværelsen av en annen spesialisert type celler kjent som eosinofiler, hvis andel øker etter eksponering for orale allergener.
Epitel assosiert med lymfoide follikler
Ileon er dekket av et enkelt epitel (et enkelt lag med celler) anordnet sylindrisk. I regionene ved siden av lymfoide follikler av peyerplatene er det imidlertid et stort antall plateepitelceller kjent som M -celler, mikroplo -celleceller eller spesialiserte membranceller.
Tilsynelatende er hovedfunksjonen til cellene ved siden av disse folliklene å fange antigener og lede dem eller overføre dem til makrofager som også er assosiert med Peyer -platene.
M -celler har ikke mikrovinger og utfører aktivt pinocytose for å få transport fra tynntarmen til underpitelvev.
Immunsystemet assosiert med slimhinnene er koblet til resten av kroppens immunsystem takket være aktivering og migrasjonskapasiteten til T -lymfocyttene til Peyer -platene, som kan nå den systemiske sirkulasjonen for å utøve immunfunksjonene sine.
Andre særegne egenskaper
Forskjellige når det.
- Betaler plater vaskulatur
I motsetning til andre lymfoide vev, for eksempel lymfeknuter, har ikke Peyers plater afferente lymfekar som transporterer lymfe mot deres "interiør". Imidlertid har de efferent drenering eller efferent lymfekar, som er i stand til å transportere lymfe utenfor lymfoide follikler.
Cellene inne i platene blir vannet av arterioler eller små blodkar som er i stand til å danne en kapillærbed drenert av vénulas av høyt endotel.
Relaterte sykdommer
Gitt den viktige funksjonen som Peyer -plater oppfyller i menneskekroppen, er det et stort antall tilknyttede patologier, blant dem følgende kan gjøres:
Crohns sykdom
Det er en inflammatorisk patologi som er preget av den tilbakevendende betennelsen i fordøyelseskanalen. Involvering av Peyers plater i denne sykdommen skyldes at de typiske lesjonene i dette antar at utløseren av adaptive immunresponser eller medfødt til bakteriefloraen.
Det kan tjene deg: smak papillaer: egenskaper, typer, beliggenhetI tillegg ser det ut til at Crohns sykdom spesielt påvirker den distale delen av ileum, akkurat der det er funnet en rik mengde peyerplater.
Vert eller "graft sykdom eller"Graft kontra vertssykdom " (GVHD)
Denne tilstanden er tydelig som en "kamp" mellom transplantasjoner eller transplantasjoner fra en pasient til en annen genetisk ikke -kompatibel.
Det antas at interaksjonen mellom bakteriefloraen og den epiteliale immunresponsen bidrar til fremkalling av de inflammatoriske signalene som bidrar til stimulering av D -donor -T -cellene, mediert av antigenet som presenterer celler fra verten av verten.
Deltakelsen av Peyers plater i denne prosessen ble anerkjent av Murai og samarbeidspartnere, som demonstrerte at disse strukturene er det anatomiske stedet der infiltrasjonen av donorens T -celler oppstår og hvor "anti -host" cytotoksiske celler dannes.
Referanser
- Bonnardel, J., Dasilva, ca., Henri, s., Tamoutouare, s., Chasson, l., Montañana-Sanchis, f.,… Lelouard, h. (2015). Medfødte og adaptive immunfunksjoner av Peyers lapp monocytt-avledede celler. Cellerapporter, elleve(5), 770-784.
- Collins, k. J., Cashman, s., Morgan, J., & Sullivan, G. C. ENTEN. (2012). Gastrointestinal immunsystem: samle mikrober i tarmen. Annals of Gastroenterology & Hepatology, 3(1), 23-37.
- Da Silva, C., Wagner, c., Bonnardel, J., Gorvel, J. P., & Lelouard, h. (2017). Peyer's Patch Mononuclear Phagocyte System i stabil tilstand og under infeksjon. Grenser i immunologi.
- Gartner, l., & Hiatt, j. (2002). Histology Atlas Text (2. utg.). Mexico d.F.: McGraw-Hill Inter-American Editors.
- Jung, c., Hugot, J., & Barreau, f. (2010). Peyers lapper: Immunsensorene i tarmen. International Journal of Inflammation, 1-12.
- Kagnoff, m., & Campbell, S. (1974). Funksjonelle egenskaper ved Peyers lapplymfoide celler. Yo. Induksjon av humoral antistoff og cellemedierte allografiske reaksjoner. Journal of Experimental Medicine, 139, 398-406.
- Keren, d. F., Holt, s. S., Collins, h. H., Gemski, p., Formell, s. B., Keren, d. F.,... formell, s. B. (1978). Rollen til Peyers lapper i den lokale immunresponsen fra kanin ileum for å leve bakterier. Journal of Immunology, 120(6), 1892-1896.
- Kindt, t., Goldsby, r., & Osborne, B. (2007). Kuby immunologi (6. utg.). Mexico d.F.: McGraw-Hill Inter-American fra Spania.
- Kogan, a. N., & von Andrian, u. H. (2008). Lymfocytthandel. I Mikrosirkulasjon (s. 449-482).
- Mayrhofer, g. (1997). Peyers lapporganogenese - Cytokines Rule, OK? MAGE, 41(5), 707-709.
- Mishra, a., Hogan, s. P., Brandt, e. B., & Rothenberg, m. OG. (2000). Peyers lapp eosinofiler: Identifisering, karakterisering og regulering ved slimhinneallergeneksponering, interleukin-5 og eotaxin. Blod, 96(4), 1538-1545.
- « Gastriske kjertler egenskaper, funksjoner, histologi
- Butia yatay -egenskaper, habitat, bruk, omsorg »