

Muskarin reseptorer struktur, typer og deres funksjoner, antagonister
- 1217
- 178
- Mathias Aas
De muskarinreseptorer De er molekyler som medierer acetylkolin (ACh) og er lokalisert i den postsynaptiske membranen til synapser der nevnte nevrotransmitter frigjøres; Hans navn kommer fra hans følsomhet for muskarinalkaloid produsert av soppen Amanita Muscaria.
I sentralnervesystemet er det flere neuronale sett hvis aksoner frigjør acetylkolin. Noen av dem ender i selve hjernen, mens de fleste utgjør motorveiene for skjelettmuskulatur eller effektorveiene til det autonome nervesystemet for kjertlene og hjertet og glatte muskler.
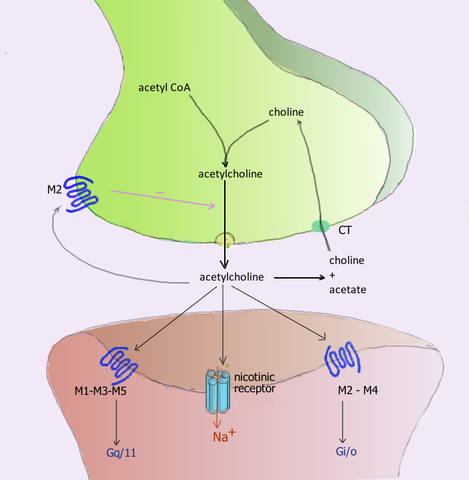
Acetylkolin -nevroreseptoren under synapsen og deres respektive reseptorer i den postsynaptiske membranen (kilde: Bruker: Pancrat [GFDL 1.2 (http: // www.gnu.Org/lisenser/old-lisens/fdl-1.2.HTML)] via Wikimedia Commons) Acetylkolin frigjort i de nevromuskulære fagforeningene i de aktive skjelettmuskelkolinergiske reseptorene kalt nikotinikk, for sin følsomhet for nikotinalkaloid, og som også finnes i synapsen gangliones i det autonome nervesystemet (SNA).
De postganglioniske nevronene i den parasympatiske inndelingen av dette systemet utøver sine funksjoner ved å frigjøre acetylkolin, som virker på de muskariniske kolinergiske reseptorene lokalisert i membranene til effektorcellene, og induserer dem elektriske modifikasjoner ved endringer av permeabilitet i deres Ionic -kanteller.
 Kjemisk struktur av acetylkolin nevrotransmitter (kilde: Neurotokeker [offentlig domene] via Wikimedia Commons) [TOC]
Kjemisk struktur av acetylkolin nevrotransmitter (kilde: Neurotokeker [offentlig domene] via Wikimedia Commons) [TOC]
Struktur
Muskarinreseptorene tilhører familien til metabotropiske reseptorer, et begrep som de reseptorene som ikke er riktig ioniske kanaler, er betegnet, men proteinstrukturer som når aktiverte trigger intracellulære metabolske prosesser som modifiserer aktiviteten til sanne kanaler.
Begrepet brukes til å skille dem fra ionotropiske reseptorer, som er sanne ioniske kanaler som åpner eller lukkes ved direkte virkning av nevrotransmitteren, som tilfellet er nikotinreseptorene som allerede.
Innenfor de metabotropiske reseptorene er muskarinikk inkludert i gruppen kjent som Grozed G de aktiverer fosfolipase C (PLC).
Muskarinreseptorer er lange omfattende membranproteiner; De har syv transmarkedssegmenter sammensatt av alfa -tilbøyeligheter, som sekvensielt krysser lipidmembranalt dobbeltlag. Inne i det cytoplasmatiske ansiktet er de assosiert med det tilsvarende G-proteinet som transduserer ligand-mottaker-interaksjonen.
Det kan tjene deg: Ebrioblasts: Utvikling, funksjoner og lagTyper muskarinreseptorer og deres funksjoner
Minst 5 typer muskarinreseptorer som er utpekt ved bruk av bokstaven M etterfulgt av et tall er identifisert, nemlig: M1, M2, M3, M4 og M5.
M1, M3 og M5 reseptorer danner M1 -familien og er preget av deres GQ- eller G11 -proteinforening, mens M2- og M4 -reseptorer er fra M2 -familien og er assosiert med GI -proteinet.
- M1 reseptorer
De finnes hovedsakelig i sentralnervesystemet, i de eksokrine kjertlene og i Nodon Autonomic System Ganglia. De er koblet til GQ -protein, som aktiverer fosfolipase C -enzymet, som konverterer til fosfatidyl inositol (PIP2) til inositol -tryposfat (IP3), som frigjør intrakellulært, og diCilglycerol (DAG), som aktiverer proteiner C COnellular, og DiCilglycerol (DAG), som aktiverer proteiner C.
- M2 -reseptorer
De finnes fundamentalt i hjertet, hovedsakelig i cellene i den kinesoaurikulære noden, som de virker ved å redusere utladningsfrekvensen, som beskrevet nedenfor.
Hjerteautomatisme
M2 -reseptorer har blitt grundigere studert ved den nodoaurikulære noden (SA) i hjertet, der automatismen som med jevne mellomrom produserer de rytmiske eksitasjoner som er ansvarlige for hjertemekanisk aktivitet, normalt manifesteres.
Cellene i den sinoaurikulære noden, etter hvert handlingspotensial (PA) som utløser en hjertesystole (sammentrekning), blir repolarisert og returnert til nivået på omtrent -70 mV. Men spenningen forblir ikke i den verdien, men lider progressiv depolarisering til et terskelnivå som utløser et nytt handlingspotensial.
Denne progressive depolariseringen skyldes spontane endringer i ioniske strømmer (I) som inkluderer: K+ (IK1) utgangsreduksjon, utseende av en oppføringstrøm på Na+ (FI) og deretter en oppføring av Ca ++ (ICAT), til det når terskelen og en annen Ca ++ (ical) strøm som er ansvarlig for handlingspotensialet utløses.
Hvis utgangen av K+ (IK1) er veldig lav og inngangsstrømmene til Na+ (hvis) og Ca ++ (ICAT) er høye, oppstår depolarisering raskere, handlingspotensial og sammentrekning oppstår tidligere, og frekvenshjerte er høyere. De motsatte modifikasjonene i disse strømningene lavere frekvens.
Kan tjene deg: cariocinesisMetabotropiske forandringer indusert av noradrenalin (sympatisk) og acetylkolin (parasympatisk) kan endre disse strømningene. AMPC aktiverer direkte IF -kanaler, proteiner A (PKA) Phosphoryila og aktiverer ICAT Ca ++ -kanaler og β GI -proteingruppen aktiverer utgangen til K+.
Muscarinic Action M2
Når acetylkolin frigjort av de postganglioniske termineringene av de hjerteskjagale fibrene (parasympatisk) binder seg til M2 -muskarinreseptorene til cellene i den sinoaurikulære noden, endrer αi -underenheten til GI -proteinet sitt GDP med GTP og skiller seg fri til BLOCK -proteinet.
ΑI -underenheten hemmer adenylciclase og reduserer produksjonen av AMPC, noe som reduserer aktiviteten til kanalene for IF og PKA. Dette siste faktum reduserer fosforylering og aktivitet av Ca ++ -kanaler for ICAT; Resultatet er en reduksjon i depolariserende strømmer.
Gruppen dannet av βy -underenhetene til GI -proteinet aktiverer en strøm av K+ Out (Ikach) som har en tendens til å motvirke inngangene til Na+ og Ca ++ og senke depolariseringshastigheten.
Fellesresultatet er en reduksjon i skråningen av spontan depolarisering og en reduksjon i hjerterytmen.
- M3 reseptorer
 M3 Muscarinic Receptor Scheme (Kilde: Takuma-SA [CC0] via Wikimedia Commons)
M3 Muscarinic Receptor Scheme (Kilde: Takuma-SA [CC0] via Wikimedia Commons) De kan finnes i den glatte muskelen (fordøyelsessystem, blære, blodkar, bronki), i noen eksokrine kjertler og i sentralnervesystemet.
GQ -protein er også koblet, og på lungenivå kan de forårsake bronkokonstriksjon, mens de virker i den vaskulære endotelfrigjøringen nitrogenoksid (NO) og forårsaker vasodilatasjon.
- M4- og M5 -reseptorer
Disse reseptorene er mindre karakterisert og studert enn de forrige. Hans tilstedeværelse i sentralnervesystemet og i noen perifere vev er blitt rapportert, men deres funksjoner er ikke tydelig etablert.
Kan tjene deg: Tata -boks: Kjennetegn og funksjonerAntagonister
Den universelle antagonisten for disse reseptorene er atropin, en alkaloid utvunnet fra planten Belladone Atropa, som binder dem med en høy affinitet, som representerer et kriterium for å skille dem fra nikotiniske reseptorer som er ufølsomme for dette molekylet.
Det er et stort antall andre antagonistiske stoffer som binder seg til de forskjellige typene muskarinreseptorer med forskjellige affiniteter. Kombinasjonen av forskjellige affinitetsverdier for noen av dem har tjent nettopp for inkludering av disse reseptorene i en eller annen av kategoriene beskrevet.
En delvis liste over andre antagonister vil omfatte: pirenzepin, metotektramin, 4-damper, hymbacin, AF-DX 384, Tripitramine, Darifenacin, PD 102807, Aq RA 741, Pfhhsid, MT3 og MT7; giftstoffer sistnevnte inneholdt i henholdsvis giftene med de grønne og svarte mambasene.
M1 -reseptorer har for eksempel høy følsomhet for pirenzepin; M2 ved tripitramin, metctramin og hybacin; M3 med 4-fuktig; M4 er veldig relatert til MT3 -toksin og også til hybacin; M5 ligner veldig på M3, men med hensyn til dem er de mindre relatert til her 741.
Referanser
- Ganong WF: Nevrotransmittere og nevromodulatorer, i: Gjennomgang av medisinsk fysiologi, 25. utg. New York, McGraw-Hill Education, 2016.
- González JC: Muskarinreseptorer rolle i moduleringen av GABAergic overføring i hippocampus. Minne for å velge legegrad. Autonome University of Madrid. 2013.
- Guyton AC, Hall JE: Rytmisk eksitasjon av hjertet, i: Lærebok for medisinsk fysiologi , 13. utg; Ac Guyton, JE Hall (eds). Philadelphia, Elsevier Inc., 2016.
- Piper HM: Herzerregung, i: Physiologie des Menschen Mite Pathophysiologie, 31. utg; RF Schmidt et al (Eds). Heidelberg, Springer Medizin Verlag, 2010.
- Schrader J, Gödeche A, Kelm M: Das Hertz, IN: Fysiologi, 6. utg; R Klinke et al (Eds). Stuttgart, Georg Thieme Verlag, 2010.
- Siegelbaum SA, Clafam DE, Schwartz JH: Modulering av synaptisk overføring: Second Messengers, In: Principles of Neural Science, 5th Ed; E Kandel et al (eds). New York, McGraw-Hill, 2013.
- « Cellular Biology History, hvilke studier, applikasjoner og konsepter
- Hvordan reproduserer planter? Aseksuell og seksuell reproduksjon »