

Reninstruktur, produksjon, sekresjon, funksjoner
- 846
- 223
- Thomas Karlsen
De Renina, Også kjent som angiotensinogease, det er en aspartylprotease som har viktige implikasjoner i elektrolytthomeostase og blodtrykkskontroll hos pattedyr.
Dette proteinet skilles ut fra nyren til sirkulasjonsstrømmen og er ansvarlig for å øke blodtrykket hos eksperimentelle dyr når nyreekstrakter blir injisert.
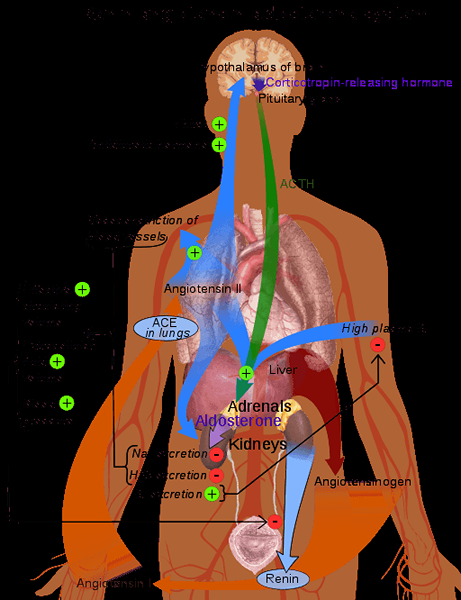
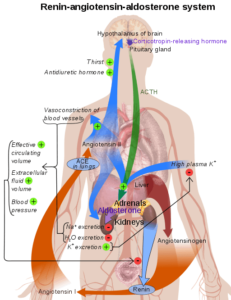 Representative ordning av renin-angiotensinsystemet i menneskekroppen (kilde: Mikael Häggström [Public Domain] via Wikimedia Commons)
Representative ordning av renin-angiotensinsystemet i menneskekroppen (kilde: Mikael Häggström [Public Domain] via Wikimedia Commons) Siden det er et stoff produsert av et stoff og utskilt til sirkulasjonen med et mål borte fra produksjonsstedet, regnes Renin som et hormon.
Hormoner kan være proteiner eller polypeptider, har steroidopprinnelse eller stammer fra aminosyret tyrosin. Renin er et hormon av proteinkarakter og dens katalytiske virkning innebærer den enzymatiske klivaje av andre proteiner (det er en protease).
Dette hormonet ble oppdaget på slutten av 1890 -tallet. Imidlertid var det ikke annet enn på slutten av 1990 at dens fysiologiske opprinnelse og molekylstruktur ble bestemt nøyaktig.
[TOC]
Struktur
Human Renin er et glykoprotein med enzymatisk aktivitet og en molekylvekt på litt over 37 kDa. Molekylet dannes av to domener atskilt med en dyp spalte som det aktive stedet er lokalisert.
Begge domenene til renin er like i rekkefølge og er hovedsakelig sammensatt av ß-belagte ark.
Ulike analyser av sekvensen av dette proteinet avslører at det har mer enn 30 basiske aminosyreavfall, inkludert flere argininer, lysiner og histidiner.
I tillegg er det kjent at det gjennom hele strukturen er hydrofobt sentrum og store hydrofile overflater som gir stabilitetsprotein i forskjellige sammenhenger.
Det aktive stedet for enzymet er lokalisert i spaltet som er dannet av de to domenene, og de essensielle aminosyrene for katalyse er to aspartinsyreavfall i posisjoner 38 og 226, og det er derfor dette er en "aspartil" -protease.
Produksjon
Reninen forekommer i det juxtaglomerulære systemet i nyren, en spesialisert struktur som finnes på kontaktstedet mellom den distale konturerte tubulus og dens opprinnelsesglomerulus.
Dette apparatet består av tre komponenter: de granulære cellene, de ekstraaglomerulære mesangiale cellene og den tette makulaen.
Tett makula
Den tette makulaen dannes av en rad med kubiske epitelceller koblet veldig kompakt som polstret røret på kontaktstedet med glomerulus, og som regnes som begynnelsen på den distale konturerte tubulus.
Kan tjene deg: Ribulosa: Karakteristikker, struktur og funksjonerMesangiale celler
Ekstraglomerulære mesangiale celler danner et trekantet område mellom den afferente arteriola, den efferente arteriola og den tette makulaen, de regnes som en forlengelse av de glomerulære mesangiale cellene. De kalles også agranulære celler.
Granulære celler
Granulære celler kalles juxtaglomerular celler og er lokalisert på veggene i afferente og efferente arterioler og i området ekstraglomerulære mesangiale celler.
Disse granulære cellene får denne kirkesamfunnet for tilstedeværelse av sekresjonsgranuler i deres cytoplasma. Granuler som inneholder renin, så vel som en forløper for renin, pro-reein, som er dannet fra pre-pr-Renine.
Pre-Pro-Renina er et prehormon som har 406 aminosyrer hos mennesker. Dette prehormonet gjennomgår en post-translasjonell proteolytisk Clivaje, som mister en sekvens på 23 avfall ved sin terminale aminoend.
Pre-Pro-Renins Cliving konverterer den til pro-Reign, 383 aminosyrer. Den bakre clivaje av en annen sekvens i den N-terminale enden av pro-reign er det som leder dannelsen av renin, en aktiv protease på 340 aminosyrer.
Både pro-rrenin og renin kan skilles ut til sirkulasjon, men veldig lite pro-reign blir omdannet til aktiv renin til dette bindevevet. Enzymene som har ansvaret for konvertering av renin renin er kjent som kalicreiner og katopsiner.
Når reninen er utskilt til sirkulasjonen, har den et halvt liv som ikke overstiger 80 minutter, og sekresjonen er sterkt regulert.
I tillegg til nyren, kan renin produseres av andre vev eller organer som testikler, eggstokker, veggene i arteriolene, binyrene, hypofysen, hjernen, fostervannet og andre.
Selv om det er aktuelt for mange dyr, viser studier som involverer fjerning av nyrene at reninaktiviteten i sirkulasjonen faller drastisk til nivåer veldig nær null.
Sekresjon
Reninens sekresjon øker med en serie stimuli som vises når volumet av den ekstracellulære væsken avtar, når blodtrykket synker eller når den sympatiske aktiviteten i nyre nerver øker.
Flere faktorer relatert til regulering av reninsekresjon er beskrevet:
- Renal perfusjonstrykket som ble påvist av baroreceptorene (strekkreseptorer) av den afferente arteriola
- Endringer i volumet og sammensetningen av væsken som når den tette makulaen
- Renal sympatiske nerveraktiviteter
- Prostaglandiner
- Det natriuretiske atriepeptidet.
Baroreceptor -mekanismen til den afferente arteriola forårsaker en reduksjon i reninsekresjon når en økning i trykket til den afferente arteriola på nivået av den juxtaglomerulære enheten oppstår. Sekresjonen øker når baroreceptoraktiviteten avtar når trykket synker.
Kan tjene deg: stroma (histologi)En annen sensor relatert til regulering av reninsekresjon finnes i den tette makulaen. Jo høyere reabsorpsjonshastighet for Na+ og Cl- og konsentrasjonen av disse elektrolyttene i væsken som når den tette makulaen, jo lavere er sekresjonen av renin og omvendt.
Økningen i nyresympatisk nerveaktivitet, samt sirkulerende katekolaminer gjennom noradrenalin frigitt i sympatiske termineringer i juxtaglomerular celler, øker reninsekresjonen.
Prostaglandiner, spesielt prostacicline.
Angiotensin II, på grunn av en negativ tilbakemeldingseffekt, hemmer reninsekresjon ved direkte effekt på granulære celler. Et annet hormon som vasopressin hemmer reninsekresjon.
Det natriuretiske atriepeptidet (PNA), som forekommer i hjertestrisk muskel, hemmer reninsekresjon.
Den ekstra effekten av alle stimulerende og hemmende faktorer er det som bestemmer reninsekresjonshastigheten. Renin skilles ut i nyreblod og forlater deretter nyrene for å sirkulere gjennom hele kroppen. Imidlertid gjenstår en liten mengde renin i nyrefluid.
Funksjoner
Renin er et enzym som i seg selv ikke har vasoaktive funksjoner. Den eneste kjente funksjonen til Renin er å kutte angiotensinogen ved aminotionenden, og generere en halshugget kalt angiotensin I.
Angiotensinogen er et glykoprotein fra gruppen av α2 globuliner syntetisert av leveren og er til stede i det sirkulerende blodet.
Siden angiotensin har jeg en veldig dårlig vasopressoraktivitet og den må behandles "nedstrøms" av en annen protease, deltar reninen i de første trinnene i blodtrykksregulering, i et system kjent som renin-angiotensin.
Angiotensin II har en veldig kort halvliv (mellom 1 og 2 minutter). Det metaboliseres raskt av flere peptidaser som fragmenterer det og noen av disse fragmentene, for eksempel angiotensin III beholder en viss vasopressoraktivitet.
De generelle funksjonene til renin -angiotensin -systemet er flere og kan oppsummeres som følger:
- Arteriolar innsnevring og økning i systolisk og diastolisk trykk. Angiotensin II er fire til åtte ganger kraftigere enn noradrenalin for denne funksjonen.
Kan tjene deg: Hyracotherium: Kjennetegn, ernæring, arter, reproduksjon- Økning i aldosteronutskillelse på grunn av direkte effekt av angiotensin II på binyrebarken. Renin-angiotensinsystemet er hovedregulatoren for aldosteronsekresjon.
- Det letter noradrenalinutskillelse på grunn av direkte effekt på sympatiske etterganglioner.
- Det påvirker mesangialcellekontraksjon, noe som reduserer den glomerulære filtreringshastigheten, og på grunn av direkte effekt på nyretubuli øker natriumreabsorpsjonen.
- På hjernenivå reduserer dette systemet følsomheten til baroreceptorrefleksen som forbedrer angiotensin II Vasopressor -effekten.
- Angiotensin II stimulerer vanninntaket ved å fremme tørstmekanismer. Øker sekresjonen av vasopressin og ACTH -hormon.
Relaterte patologier
Renin-angiotensinsystemet har derfor en viktig rolle i hypertensive patologier, spesielt de med renal opprinnelse.
Slik genererer innsnevringen av en av nyrearteriene vedvarende hypertensjon som kan reverseres hvis den iskemiske (mangelfulle) nyren blir trukket ut eller renal arteriell innsnevring frigjøres i tid.
En økning i reninproduksjon er generelt assosiert med den ensidige innsnevringen av nyrearterien som forbinder en av nyrene, noe som forårsaker hypertensjon. Denne kliniske tilstanden kan skyldes medfødte defekter eller andre avvik i nyresirkulasjonen.
Den farmakologiske manipulasjonen av dette systemet, i tillegg til bruk av blokkering av angiotensin II -reseptorer, er de grunnleggende verktøyene for behandling av arteriell hypertensjon.
Arteriell hypertensjon er en stille og progressiv sykdom som rammer en stor del av verdens befolkning, spesielt 50 år gammel voksne.
Referanser
- Akahane, k., Umeyama, h., Nakagawa, s., Moriguchi, i., Hirose, s., Iizuka, k., & Murakami, J. (1985). Tredimensjonal struktur av menneskelig renin. Hypertensjon, 7(1), 3-12.
- Davis, J., & Freeman, R. (1976). Mekanismer som regulerer reninavvik. Fysiologiske anmeldelser, 56(1), 1-56.
- Guyton, a., & Hall, J. (2006). Lærebok for medisinsk fysiologi (11. utg.). Elsevier Inc.
- Hackenthal, e., Paul, m., Ganten, d., & Taugner, r. (1990). Morfologi, fysiologi og molekylærbiologi av reninhemmelighet. Fysiologiske anmeldelser, 70(4), 1067-1116.
- Morris, f. (1992). Biologi av reninmolekylær. I: Gen- og proteinstruktur, syntese og prosessering. Journal of Hypertensjon, 10, 209-214.
- Murray, r., Bender, d., Botham, k., Kennelly, p., Rodwell, v., & Weil, P. (2009). Harpers illustrerte biokjemi (28. utg.). McGraw-Hill Medical.
- West, J. (1998). Fysiologisk grunnlag for medisinsk praksis (12ava ed.). Mexico d.F.: Pan -American Medical Editorial.