

Spleising (genetikk)
- 1359
- 314
- Prof. Theodor Gran
RNA ledddiagram Hva er skjøting?
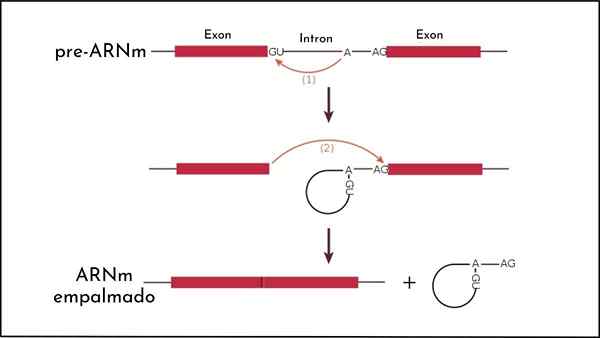
Han Spleising O skjæreprosess og RNA -spleising er et molekylært fenomen som oppstår i eukaryote organismer etter transkripsjonen av DNA til RNA og involverer eliminering av intronene i et gen, og beholder eksonene. Anses som grunnleggende i genuttrykk.
Det skjer gjennom hendelser for å eliminere fosfodisterkobling mellom eksoner og introner og den påfølgende Bond -unionen mellom eksoner.
Spleising skjer i alle typer RNA; Imidlertid er det mer relevant i Messenger RNA -molekylet. Det kan også forekomme i DNA- og proteinmolekyler.
På tidspunktet for montering av eksonene lider de en ordning eller noen endringshastighet. Denne hendelsen er kjent som alternativ spleising og har viktige biologiske konsekvenser.
Hva er skjøting?
Et gen er en DNA -sekvens med nødvendig informasjon for å uttrykke en fenotype. Genbegrepet er ikke strengt begrenset til DNA -sekvenser som uttrykkes som proteiner.
Den sentrale "dogmen" av biologi involverer prosessen med transkripsjon av DNA til et mellomliggende molekyl, Messenger RNA. Dette oversettes igjen til proteiner ved hjelp av ribosomer.
I imidlertid eukaryote organismer blir disse lange sekvensene av gener avbrutt av en type sekvenser som ikke er nødvendige for det aktuelle genet: introner. For at Messenger RNA kan oversettes effektivt, må disse intronene elimineres.
RNA -spleising er en mekanisme som involverer flere kjemiske reaksjoner, som brukes til å fjerne elementer som avbryter sekvensen til et visst gen. Elementene som er bevart kalles eksoner.
Det kan tjene deg: hollandisk arv: egenskaper, genereringsfunksjoner, degenerasjonDer spleisingen oppstår?
Espliceosoma er et stort protein -naturkompleks som er ansvarlig for å katalysere trinnene for skjøting. Den består av fem typer små-nukleært RNA, kalt U1, U2, U4, U5 og U6, i tillegg til en serie proteiner.
Det spekuleres i at forklaringen deltar i foldingen av pre-ARNM for å justere det riktig med de to regionene der skjøteprosessen vil oppstå.
Dette komplekset er i stand til å gjenkjenne konsensus -sekvensen som de fleste introner har om endene 5 'og 3'. Det skal bemerkes at det er funnet gener i metazoa som ikke har disse sekvensene og bruker en annen liten-kjernefysisk RNA-gruppe for gjenkjennelse.
Spleisende typer
I litteratur blir begrepet spleising vanligvis brukt på prosessen som involverer messenger -RNA. Imidlertid er det forskjellige skjøteprosesser som oppstår i andre viktige biomolekyler.
Proteiner kan også eksperimentere skjøting, i dette tilfellet er det en sekvens av aminosyrer som fjernes fra molekylet.
Det eliminerte fragmentet kalles "Intenlena". Denne prosessen skjer naturlig i organismer. Molekylærbiologi har klart å lage forskjellige teknikker ved å bruke dette prinsippet som involverer proteinkanipulering.
Tilsvarende forekommer skjøting også på DNA -nivå. Dermed er to DNA -molekyler som tidligere var atskilt, i stand til å bli sammen med kovalente bindinger.
Typer RNA -skjøting
På den annen side, avhengig av type RNA, er det forskjellige kjemiske strategier der genet kan bli kvitt introner.
Spesielt spleising før ARNM er en komplisert prosess, siden den innebærer en serie trinn katalysert av espliceosomet. Kjemisk oppstår prosessen på grunn av transcessterifiseringsreaksjoner.
Kan tjene deg: dominerende allel: egenskaper og eksemplerI gjær, for eksempel, begynner prosessen med brudd på region 5 'på gjenkjennelsesstedet, er intron-exon "loop" dannet av en 2'-5' fosfodiéster-kobling. Prosessen fortsetter med dannelsen av et gap i region 3 ', og til slutt oppstår foreningen av de to eksonene.
Noen av intronene som avbryter kjernefysiske og mitokondriergener kan utføre skjøting uten behov for enzymer eller energi, men ved hjelp av transcessterifiseringsreaksjoner. Nevnte fenomen ble observert i kroppen Tetrahymena Thermophila.
I kontrast tilhører de fleste kjernefysiske gener gruppen av introner som trenger maskiner som katalogiserer eliminasjonsprosessen.
Alternativ spleising
Hos mennesker er det rapportert at det er rundt 90.000 forskjellige proteiner, og trodde tidligere at det skulle være et identisk antall gener.
Med ankomsten av nye teknologier og Human Genome -prosjektet, kan det konkluderes med at vi bare har omtrent 25.000 gener. Så hvordan er det mulig at vi har så mange proteiner?
Ekonene er kanskje ikke samlet i samme rekkefølge som de ble transkribert til RNA, men er fikset ved å etablere nye kombinasjoner.
Dette fenomenet er kjent som alternativ spleising. Av denne grunn kan et enkelt transkribert gen produsere mer enn en type protein.
Denne uoverensstemmelsen mellom antall proteiner og antall gener ble belyst i 1978 av forsker Gilbert, og etterlot det tradisjonelle konseptet "av et gen er det et protein".
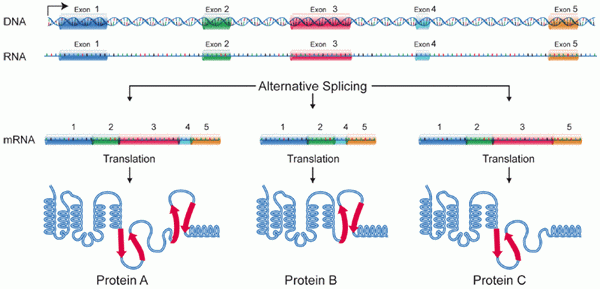 Alternativt spleisediagram. Kilde: National Genome Research Institute, Wikimedia Commons
Alternativt spleisediagram. Kilde: National Genome Research Institute, Wikimedia Commons Funksjoner
For Kelemen og samarbeidspartnere (2013), "en av funksjonene til denne hendelsen er.
Kan tjene deg: multifaktoriell arvI følge disse forfatterne er "alternativ spleising ansvarlig for å regulere plasseringen av proteiner, deres enzymatiske egenskaper og deres interaksjon med ligander". Det har også vært relatert til prosessene med celledifferensiering og utvikling av organismer.
I lys av evolusjonen ser det ut til å være en viktig mekanisme for endring, siden en høy andel av høyere eukaryote organismer som lider av høye alternative skjøtehendelser er funnet. I tillegg til å spille en viktig rolle i differensieringen av arter og i genomets utvikling.
Alternativ spleising og kreft
Det er bevis på at enhver feil i disse prosessene kan føre til unormal funksjon av cellen, og gi alvorlige konsekvenser for individet. Innenfor disse potensielle patologiene skiller kreft seg ut.
Det er grunnen til at alternativ spleising er blitt foreslått som en ny biologisk markør for disse unormale forholdene i celler.
På samme måte, hvis grunnlaget for mekanismen som sykdommen oppstår i dybden oppnås, kan løsninger foreslås for dem.
Referanser
- Berg, J. M., STRYER, L., & Tymoczko, J. L. (2007). Biokjemi. Jeg snudde meg.
- Av conti, l., Baralle, m., & Buratti, og. (2013). Exon og intron definisjon i pre -mrna spleising. Wiley tverrfaglige anmeldelser: RNA, 4(1), 49-60.
- Kelemen, eller., Convertini, s., Zhang, Z., Wen, og., Shen, m., Falaleeva, m., & Stamm, S. (2013). Funksjon av alternativ spleising. Gen, 514(1), 1-30.
- Lamond, a. (1993). Spliceosomet. Bioessays, 15(9), 595-603.
- Vila-Perelló, m., & Muir, t. W. (2010). Biologiske anvendelser av proteinskjøting. Celle, 143(2), 191-200.