

Bein stoffegenskaper, struktur, dannelse og vekst
- 1207
- 192
- Jonathan Moe
Han vevd bein er den som komponerer beinene. Benet, sammen med emalje og dentin, er de tøffeste stoffene i dyrenes kropp. Benene utgjør strukturene som beskytter de vitale organene: Hjernen er beskyttet av hodeskallen, ryggmargen ved ryggraden, og hjertet og lungene er gjennom ribbeholderen.
Benene fungerer også som "spaker" for musklene som settes inn i dem, og multipliserer kraften som disse musklene genererer under utførelsen av bevegelsene. Stivheten gitt av beinet tillater bevegelse og støtte av belastningene mot tyngdekraften.
Benvevsceller (kilde: OpenStax College [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) Benet er et dynamisk levende stoff som stadig endres, og disse endringene stimuleres av trykket og spenningene som dette vevet blir utsatt for. For eksempel stimulerer trykk resorpsjon (ødeleggelse) og spenning kan stimulere ny beindannelse.
Benene utgjør hovedforekomsten av kalsium og fosfor av organisme: nesten 99% av det totale kalsiumet i menneskekroppen lagres i beinvevet. Total beinmasse varierer gjennom et dyrs levetid. I løpet av vekstfasen overstiger beindannelse resorpsjonen (ødeleggelse) og skjelettet vokser og utvikler seg.
Opprinnelig øker lengden og deretter dens tykkelse, og når det maksimum mellom 20 og 30 år hos mennesker. Hos den voksne (opptil omtrent 50 år) er det en balanse mellom formasjon og beinresorpsjon.
Denne balansen er gitt av en erstatningsprosess som er kjent som "beinoppussing" og som påvirker per år fra 10% til 12% av den totale beinmassen. Deretter begynner en degenerativ prosess der resorpsjonen overstiger dannelsen og beinmassen synker sakte.
[TOC]
Egenskaper og struktur
Benet har et sentralt hulrom som kalles kjernehulrom, som huser benmargen, et hematopoietisk vev, det vil si et blodcelle som danner vev. Disse strukturene er dekket av periosteum, med unntak av områdene som tilsvarer synoviale leddene.
Periosteium har et ytre lag med fibrøst tett bindevev og et indre lag med osteogene celler, som er beindannende celler eller osteoprogenceller.
Den sentrale delen av beinet blir polstret av en tynn og spesialisert bindevevscelle monolay. Endostio har osteoprogen- og osteoblastceller. Benet dermed polstret, har cellene sine integrert i en matrise [F1] [F2] ekstracellulær forkalket.
Osteoprogenitorceller er forskjellige i osteoblaster og har ansvaret for sekresjon av beinmatrisen. Når de er omgitt av matrise, blir disse cellene inaktivert og osteocytters navn inaktiveres.
Rommene okkupert av osteocytter i matrisen kalles laguner.
90% av den organiske matrisen er dannet av type I kollagenfibre, et strukturelt protein også til stede i sener og hud, og resten er et homogent geléstoff som kalles grunnleggende stoff.
Kompakt bein og svampaktig bein
Kollagenfibrene til matrisen er anordnet i store bjelker, og i det kompakte beinet danner disse fibrene konsentriske lag rundt kanalene som blodkarene og nervefibrene (Havers Canals) kjører (Havers). Disse lagene danner sylindere kjent som "osteoner".
Hver osteon er avgrenset av en sementlinje dannet av grunnleggende stoff forkalket med få kollagenfibre og gir næring til karene som er i hover -kanalene.
I det svampete beinet dannes store plakk eller spikler og cellene næres ved diffusjon av den ekstracellulære væsken i beinet til trabeculae.
De uorganiske komponentene i matrisen utgjør rundt 65% av beinets tørrvekt og er hovedsakelig dannet av kalsium og fosfor, i tillegg til noen elementer som natrium, kalium, magnesium, sitrat og bikarbonat, blant andre natrium, kalium, magnesium, sitrat og bikarbonat,.
Kan tjene deg: Merychippus: Kjennetegn, reproduksjon, ernæring, taksonomiKalsium og fosfor danner hydroksyapatittkrystaller [CA10 (PO4) 6 (OH) 2]. Kalsiumfosfat finnes også i amorf form.
Hydroksyapatittkrystaller er ordnet i orden.
Beindannelse og vekst
Skullbenene dannes av en prosess kjent som "intramembranøs ossifikasjon". I stedet modelleres lange bein først i brusk og deretter transformert til bein ved ossifisering, som begynner i diaphysen av beinet og kalles "endokondral ossifikasjon".
De fleste flate bein utvikler seg og vokser ved intramembranøs beindannelse og ossifikasjon. Denne prosessen skjer i det veldig vaskulariserte mesenkymale vevet, der mesenkymale celler er forskjellige i osteoblaster som begynner å produsere beinmatrise.
Slik dannes et nettverk av spikler og trabeculae, hvis overflater er befolket av osteoblaster. Disse innledende osteogeneseområdene kalles primær ossifikasjonssenter. Dette danner det primære beinet med tilfeldig orienterte kollagenfibre.
Da oppstår forkalkningen og osteoblastene fanget i matrisen osteocytter, hvis utvidelser gir opphav til kanaler. Ettersom trabekulære nettverk dannes som en svamp, gir vaskulært bindevev opphav til benmargen.
Tilsetningen av perifere trabeculae øker beinstørrelsen. I Occipital -beinet (et kranialt bein i den bakre sonen) er det flere ossifikasjonssentre som smelter sammen for å danne et enkelt bein.
Hos nyfødte er rørleggerarbeid mellom fronten og parietalbenene ossifiseringssoner som ennå ikke er slått sammen.
Kompakt beindannelse
De mesenkimatøse vevsregionene som gjenstår uten å beregne i indre og ytre deler vil danne periosteum og endostio. De svampete beinområdene umiddelbart til periosteum og duramadre vil bli kompakt bein og danne det indre og ytre bordet til det flate beinet.
Under vekst, i lange bein, skilles områder som er spesialisert i epifyser fra diaphysen med en veldig aktiv bruskplate som kalles epifysisk plate.
Benlengden øker i den grad denne platen avsetter nytt bein i hver ende av diaphysen. Størrelsen på epifysale platen er proporsjonal med veksthastigheten og påvirkes av flere hormoner.
Regulering
Blant hormonene som modulerer denne platen er veksthormonet (GH) frigitt av den forrige hypofyseveksten som ligner på type I insulin (IGF-I) produsert av leveren.
Mens den mitotiske aktivitetshastigheten i spredningssonen ligner på benresorpsjonshastigheten til området, forblir størrelsen på epifysealplaten konstant og beinet fortsetter å vokse.
Etter 20 år reduseres mitotisk aktivitet og ossifikasjonssonen når brusksonen, og blir sammen med kjernehulene i diaphyse og epifyser.
Den langsgående veksten av beinet ender når den epifysiske lukking skjer, det vil si når diaphysen er sammen med epifyser. Epifysale lukking følger en ordnet midlertidig sekvens som ender med den siste nedleggelsen etter pubertet.
Breddeveksten av det lange beinet produseres av apostal vekst, som er et produkt av differensieringen av osteoprogenitorcellene i det indre laget av periosteum i osteoblaster som utskiller beinmatrise til de subperiostiske områdene i diaphysen.
Benombygging
Gjennom et menneske levetid erstattes beinet kontinuerlig gjennom dannelse og resorpsjonsprosesser, det vil si ødeleggelse av det gamle beinet og ny beindannelse.
Det kan tjene deg: Sinaloa Flora og Fauna: Mer vanlige dyr og planterHos spedbarn lider kalsium en 100% årlig erstatning, mens det hos voksne bare er 18% per år. Disse resorpsjons- og dannelses- eller erstatningsprosessene kalles beinoppussing.
Ombyggingen begynner med handlingen fra osteoklastene som ødelegger beinet og etterlater noen kløfter som deretter blir invadert av osteoblastene. Disse osteoblastene utskiller matrisen som deretter blir ossifisert og gir opphav til det nye beinet. Denne syklusen krever i gjennomsnitt mer enn 100 dager.
På et gitt tidspunkt er mer eller mindre 5% av hele beinmassen til skjelettet i ferd med å ombygging. Dette innebærer deltakelse av omtrent to millioner ombyggingsenheter.
Forskjeller i ombygging av det kompakte og svampete beinet
Den årlige kompakte beinoppussingshastigheten er 4 % og svampaktig bein er 20 %.
Forskjellen mellom ombyggingshastighetene for de to beintypene skyldes mest sannsynlig at det svampete beinet er i kontakt med benmargen og er direkte påvirket av cellene med paracrinaktivitet av nevnte medulla.
Osteoprogencellene i de kompakte beinene finnes tvert imot i de bjørniske kanalene og i de indre lagene i periosteum, langt fra cellene i benmargen og er avhengige, for begynnelsen av ombyggingen, av hormonene som ankommer med blod.
Mange er hormonelle og proteinfaktorer som er involvert i aktiviteten til osteoblaster og osteoklaster i beinombygging, men det har ikke vært mulig å tydelig belyse funksjonen til hver.
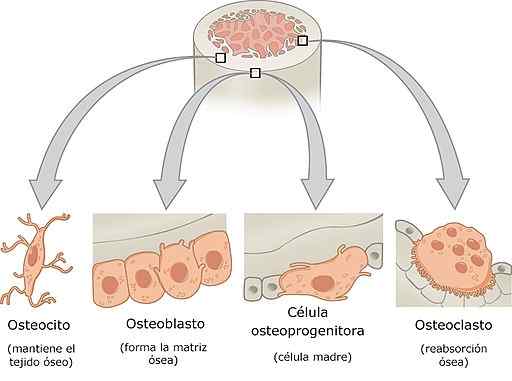
Beinceller
-Typer beinceller og deres egenskaper
Benceller er osteoprogenitorceller, osteoblaster, osteocytter og osteoklaster. Hver av disse cellene har spesielle funksjoner i beinfysiologi og har veldig differensierte histologiske egenskaper.
Osteoblaster, osteocytter og osteoklaster, sammen, danner beinmodellenheten.
Osteoprogen eller osteogene celler
Disse cellene finnes i det indre laget av periosteum og i endostio. De stammer fra det embryonale mesenkym og kan gi opphav til differensiering, til osteoblaster. Under visse stressforhold kan de også differensiere i kondrogene celler.
De er spindelformet celler med en oval kjerne, knapp cytoplasma, med lite grov endoplasmatisk retikulum (RER) og en dårlig utviklet Golgi -enhet. De har rikelig ribosomer og er veldig aktive i beinvekstperioden.
Osteoblastene
Osteoklaster er celler avledet fra osteogene celler. De er ansvarlige for å syntetisere den organiske matrisen til beinet, det vil si kollagen, proteoglykaner og glykoproteiner. De er ordnet i lag lagt på benoverflaten.
Kjernen er plassert på motsatt side av den sekretoriske delen rik på vesikler. De har rikelig RER og en velutviklet Golgi -enhet. De har korte anslag eller utvidelser som tar kontakt med andre nabolandende osteoblaster. Andre lange utvidelser kobler dem til osteocytter.
Ettersom osteoblastene utskiller foreldre.
Selv om det meste av beinmatrisen blir forkalket, er det rundt hver osteoblast og til og med hver osteocyt.
I cellemembranen til osteoblaster er det forskjellige typer reseptorer. Av disse reseptorene er det viktigste mottakeren for parathyreoidehormonet (PTH), som stimulerer sekresjonen av en stimulerende osteoklastfaktor som fremmer benresorpsjon.
Osteoblaster kan også utskille enzymer som er i stand til å fjerne osteoid og dermed sette osteoklaster med den forkalkede beinoverflaten for å starte resorpsjonen.
Osteocytter
Dette er celler avledet fra inaktive osteoblaster og kalles modne beinceller. De er plassert i de nevnte lugs av den forkalkede beinmatrisen. Det er mellom 20.000 til 30.000 osteocytter per kubikk millimeter bein.
Kan tjene deg: fosfatidylkolin: syntese, struktur, funksjoner, egenskaperFra lagunene utstråler osteocyttene cytoplasmatiske utvidelser som forener dem med hverandre, og danner interstitiumforeninger som ioner og små molekyler kan byttes mellom cellene mellom cellene.
Osteocytter er flate celler, med flate og få cytoplasmatiske organeller kjerner. De er i stand til å utskille stoffer mot mekaniske stimuli som forårsaker beinspenning (Transduction Mechano).
Plassen som omgir osteocyttene i lagunene kalles det periosteocytiske rommet og er fullt av ekstracellulær væske i den ikke -kalkifiserte matrisen. Det anslås at overflatearealet til periosteocyttveggene er omtrent 5000m2 og som huser et volum på omtrent 1,3 liter ekstracellulær væske.
Denne væsken blir utsatt for omtrent 20 g utskiftbart kalsium som kan reabsorberes i den sirkulasjonsstrømmen fra veggene i disse områdene, noe som bidrar til å opprettholde blodfigurene til kalsium.
Osteoklaster
Disse cellene stammer fra de samme stamceller som vevsmakrofager og sirkulerende monocytter; Disse finnes i benmargen og er stamfaderceller fra granulocytter og makrofager (GM-CFU).
Myitosen til disse stamcellene stimuleres av de stimulerende faktorene til makrofagkolonier og i nærvær av bein, fusjonerer disse forløperne og danner multinucleated celler.
En osteoklast er en stor, multinucleated og mobilcelle. Den måler omtrent 150μm i diameter og kan ha opptil 50 kjerner. Den har et basalområde der kjernene og organellene ligger, en penselkant i kontakt med det forkalkede beinet, klare perifere områder til børstekanten og et vesikulært område.
Hovedfunksjonen til disse cellene er benresorpsjonen. Når de utøver sin funksjon, lider de apoptose (programmert celledød) og dør. For å sette i gang benresorpsjonsprosessen, fester osteoklasten til beinet gjennom omfattende -kalt proteiner.
Deretter beveger protoner bomber som er H+-avhengige ATASAS, fra endosomene til det indre av membranen på en børstekant og surifiserte mediet til pH faller omtrent 4.
Hydroksyapatitt oppløses til slike pH- og kollagenfibre blir nedbrutt av syreproteaser også utskilt av disse cellene. Sluttproduktene av fordøyelsen av hydroksyapatitt og kollagen blir endocytet i osteoklasten og frigjøres deretter til den interstitielle væsken som deretter blir eliminert av urinen.
Beinvevstyper (beintyper)
Som du kanskje har lagt merke til i teksten, er det to typer beinvev, nemlig: det kompakte eller kortikale beinet og det trabecular eller svampete beinet.
Den første utgjør 80% av den totale beinmassen og finnes i diaphysen av de lange beinene, som er de rørformede delene som er anordnet mellom de to endene (epifysene) i disse beinene.
Den andre beintypen er typisk for beinene i det aksiale skjelettet, for eksempel ryggvirvlene, beinene i hodeskallen og bekkenet og ribbeina. Det ligger også i midten av lange bein. Den danner 20% av den totale beinmassen og er av vital betydning for reguleringen av kalsiummetabolisme.
Referanser
- Berne, r., & LEVY, M. (1990). Fysiologi. Mosby; International Ed Edition.
- Di fiore, m. (1976). Normal histologi atlas (2. utg.). Buenos Aires, Argentina: The Editorial Athenaeum.
- Doubek, r. W. (1950). Histologi med høyt avkastning (2. utg.). Philadelphia, Pennsylvania: Lippinott Williams & Wilkins.
- Fox, s. Yo. (2006). Menneskelig fysiologi (9. utg.). New York, USA: McGraw-Hill Press.
- Gartner, l., & Hiatt, j. (2002). Histology Atlas Text (2. utg.). Mexico d.F.: McGraw-Hill Inter-American Editors.
- Guyton, a., & Hall, J. (2006). Lærebok for medisinsk fysiologi (11. utg.). Elsevier Inc.
- Johnson, k. (1991). Histologi og cellebiologi (2. utg.). Baltimore, Maryland: The National Medical Series for Independent Study.
- Ross, m., & Pawlina, w. (2006). Histologi. En tekst og atlas med korrelert celle- og molekylærbiologi (5. utg.). Lippinott Williams & Wilkins.
- « Kompakte beinfunksjoner, struktur, funksjoner
- Jernoksid (II) struktur, nomenklatur, egenskaper, bruk »