

Topoisomerase hva er, egenskaper, funksjoner, typer
- 4739
- 1222
- Anders Mathisen
De Topoisomerase De er en type isomeraft -enzymer som modifiserer topologien til deoksyribonukleinsyre (DNA), og genererer både deres avvikling og dens krølling og overlapping.
Disse enzymene har en spesifikk rolle i lettelse av vridningsspenning i DNA slik at slike viktige prosesser kan oppstå, transkripsjonen av DNA i messenger ribonukleinsyre (RNAM) og rekombinering av DNA.
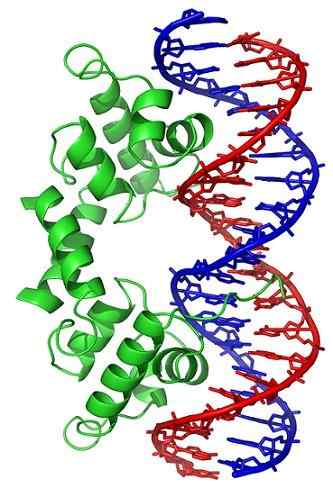
 Topoisomerase II, Wikimedia Commons
Topoisomerase II, Wikimedia Commons Topoisomerase -enzymer er til stede i både eukaryote celler og prokaryote celler. Eksistensen ble spådd av forskerne Watson og Crick, når de evaluerte begrensningene som DNA -strukturen presenterte for å gi tilgang til informasjonen deres (lagret i nukleotidsekvensen).
For å forstå funksjonene til topoisomerasen, bør det vurderes at DNA har en stabil dobbel propellstruktur, med kjedene rullet den ene på den andre.
Disse lineære kjedene er sammensatt av 2-dechexiribosas koblet med 5'-3 'fosfodi-foser.
Den topologiske studien av DNA -molekyler har vist at disse kan anta flere konformasjoner avhengig av torsjonsspenningen: fra en avslappet tilstand, til forskjellige påmeldingstilstander som tillater deres komprimering.
DNA -molekyler med forskjellige konformasjoner kalles topoisomerer. Dermed kan vi konkludere med at topoisomeraser I og II, kan øke eller redusere spenningen av torsjon av DNA -molekyler, og danne deres forskjellige topoisomerer.
Blant de mulige DNA -topoisomerer er den vanligste konformasjonen av Superstrane, som er veldig kompakt. Imidlertid må DNA -dobbeltpropell også være avviklet av topoisomeraser under flere molekylære prosesser.
Kjennetegn
Mekanisme for generell handling
Noen topoisomeraser kan bare slappe av negativ overskriving av DNA, eller begge overstrans av DNA: positiv og negativ.
Hvis det sirkulære dynamiske DNA slapper av på sin langsgående akse og en Levogira -retur (i retning av klokken nåler), sies det at det er negativt overlappende. Hvis returen er dextrogyral (i strid med klokken nåler), er den positivt overveldet.
 DNA sirkulær sirkulær høyttaler negativt, avslappet og positivt overveldet. Kilde: FDARDEL [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons
DNA sirkulær sirkulær høyttaler negativt, avslappet og positivt overveldet. Kilde: FDARDEL [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons I utgangspunktet kan topoisomeraser:
-Lette passering av en DNA -streng gjennom et kutt i motsatt streng (topoisomerase type I).
-Tilrettelegge for passering av en komplett dobbel propell gjennom en splittelse i seg selv, eller gjennom en splittelse i en annen dobbel propell (topoisomerase type II).
Oppsummert virker topoisomerase gjennom splitting av fosfodiéster -koblinger, i en eller de to trådene som utgjør DNA. Deretter modifiserer de den rullende tilstanden til strengene til en dobbel propell (molemomerase topoisomerase) eller to doble propeller (molemomerase II), for endelig å binde igjen eller binde de delte endene.
Topoisomerase og cellesyklus
Selv om topoisomerasen I er et enzym som har den største aktiviteten under fase S (DNA -syntese), anses det ikke å være avhengig av en cellesyklusfase.
Kan tjene deg: Saponifiable lipider: Kjennetegn, struktur, funksjoner, eksemplerMens topoisomerase II -aktivitet er mer aktiv under den logaritmiske fasen av cellevekst og raske vekstceller.
Funksjoner
Endringen av genene som koder. Blant prosessene der topoisomerase deltar, er:
Kompakt lagring av genetisk materiale
Topoisomeraser letter lagring av genetisk informasjonskompakte, fordi de genererer rullende og overlapping av DNA, slik at en stor mengde informasjon kan være i et relativt lite volum.
Tilgang til genetisk informasjon
Hvis det ikke er noen topoisomerase og deres unike egenskaper, ville tilgang til informasjon som er lagret i DNA være umulig. Dette skyldes det faktum at topoisomerase med jevne mellomrom frigjør spenningen på grunn av torsjon som genereres i den doble DNA -propellen, under deres avvikling, i replikerings-, transkripsjons- og rekombinasjonsprosesser.
Hvis spenningen på grunn av torsjon som genereres under disse prosessene ikke frigjøres, kan et mangelfullt genuttrykk oppstå, avbrudd i sirkulært DNA eller kromosom, til og med å produsere celledød.
Regulering av genuttrykk
Konformasjonsendringene (i den tre -dimensjonale strukturen) av DNA -molekylet, utsetter spesifikke regioner, som kan samhandle med DNA -proteiner. Disse proteinene har en regulatorisk funksjon av genetisk uttrykk (positivt eller negativt).
Genetisk ekspresjonsreguleringsprotein. Wikimedia Commons Dermed påvirker DNAs rullende tilstand, generert av virkningen av topoisomeraser, reguleringen av genetisk uttrykk.
Spesialiteter av topoisomerase II
Topoisomerase II er nødvendig for montering av kromatider, kondensasjon og kromosomer og segregering av DNA -døtre under myitose under myitose.
Dette enzymet er også et strukturelt protein, og en av hovedbestanddelene i cellekjernematrisen under grensesnittet.
Typer topoisomerase
Det er to hovedtyper av topoisomeraser avhengig av om de er i stand til å dele ut en eller to DNA -kjeder.
-Topoisomerase type I
Monomer
Topoisomerase type I er monomerer som lindrer negativ og positiv overskriving, som produseres ved bevegelse av gaffelen under transkripsjon, og under replikasjons- og genrekombinasjonsprosesser.
Topoisomerases av type I kan deles inn i type 1A og type 1B. De sistnevnte er de som finnes hos mennesker, og er ansvarlige for å slappe av DNA.
Tyrosin på sitt aktive sted
Topoisomerase 1B (TOP1B) består av 765 aminosyrer delt inn i 4 spesifikke domener. Et av disse domenene har et høyt bevart område som inneholder det aktive stedet med tyrosin (Tyr7233). All topoisomerase som er til stede på sitt aktive sted en tyrosin med en grunnleggende rolle i hele katalytiske prosessen.
Virkningsmekanismen
Tyrosinet til det aktive stedet danner en kovalent binding med 3'-fosfatenden av DNA-kjeden, kutter den og holder den sammen med enzymet, mens en annen DNA-streng passerer gjennom splittelsen.
Det kan tjene deg: flora og fauna av den ecuadorianske Sierra: Representative arterPassasjen av den andre DNA -strengen gjennom den delte strengen oppnås takket være en konformasjonstransformasjon av enzymet, som produserer åpningen av dobbelthelixen til DNA.
Deretter går topoisomerasen I tilbake til den første konformasjonen og ligater igjen endene splittet. Dette skjer på grunn av en omvendt prosess for brudd på DNA -kjeden, i det katalytiske stedet for enzymet. Endelig frigjør topoisomerasen DNA -strengen.
DNA -ligeringshastigheten er større enn delingshastigheten, noe som sikrer molekylets stabilitet og genomets integritet.
Oppsummert er type I topoisomerase:
- Splittelsen av en streng.
- Passasjen til den andre strengen gjennom splittelsen.
- Ligering av endene splittet.
-Type II topoisomerase
Mørkere
Topoisomeraser av type II er Dimérica -enzymer, som er delt.
MG -avhengig++ og ATP
Disse enzymene trenger magnesium (mg++) Og de trenger også energien som kommer fra brudd på ATP Triffosphate Link, som drar nytte av takk til en ATPase.
To aktive steder med tyrosin
Menneskelige topoisomeraser ligner veldig på gjæren (Saccharomyces cerevisiae), som består av to monomerer (underfragmenter A og B). Hver monomer presenterer et ATPASA -domene, og i et subfragment det aktive stedet tyrosin 782, som DNA kan sammenføyes med. Derfor kan to DNA -tråder kobles til topoisomerase II.
Virkningsmekanismen
Handlingsmekanismen til topoisomerase II er lik den som er beskrevet for topoisomease I, med tanke på at to DNA -kjeder er delt og ikke bare en.
I det aktive stedet for topoisomerase II er det stabilisert (gjennom en kovalent forening med tyrosin) et fragment av Dobbelt propell av DNA, kalt "Fragment G". Dette fragmentet er delt og holdes sammen til det aktive stedet av kovalente bindinger.
Deretter tillater enzymet et annet DNA -fragment, kalt "Fragment T", å passere gjennom "G" -splittfragmentet, takket være en konformasjonsendring av enzymet, som er avhengig av ATP -hydrolyse.
Topoisomerase II Liga De to endene av "G -fragmentet" og til slutt gjenoppretter sin opprinnelige tilstand, og slipper fragmentet "G". Deretter slapper DNA torsjonsspenning, slik at replikasjons- og transkripsjonsprosesser kan oppstå.
-Menneskelig topoisomerase
Det menneskelige genomet har fem topoisomerase: TOP1, TOP3a, TOP3β (av type I); og Top2α, Top2β (type II). De mest relevante humane topoisomerasene er TOP1 (topoisomerase type IB) og 2a (topoisomerase type II).
Topoisomerase -hemmere
-Topoisomerase som et kjemisk angrepsmål
Fordi prosessene katalysert av topoisomeraser er nødvendige for overlevelse av celler, er disse enzymene et godt angrephvit for å påvirke ondartede celler. Derfor anses topoisomerase som viktige i behandlingen av mange menneskelige sykdommer.
Legemidlene som interagerer med topoisomerase er for tiden veldig studert som kjemoterapeutiske stoffer mot kreftceller (i forskjellige organer i kroppen) og patogene mikroorganismer.
Kan tjene deg: glykogen: struktur, syntese, nedbrytning, funksjoner-Typer hemming
De hemmende medikamentene til topoisomerase -aktivitet kan:
- DNA -interkalat.
- Påvirke topoisomeraseenzymet.
- Interkalat til et molekyl nær det aktive stedet for enzymet mens DNA-topoisomerasekomplekset er stabilisert.
Stabiliseringen av det forbigående komplekset som dannes av foreningen av DNA til tyrosinet til det katalytiske stedet for enzymet, forhindrer foreningen av de delte fragmentene, noe som kan føre til celledød.
-Legemidler topoisomerase -hemmere
Blant forbindelsene som hemmer topoisomerase, er følgende.
Antitumorantibiotika
Antibiotika brukes mot kreft, siden de forhindrer vekst av tumorceller, og forstyrrer vanligvis DNA. De kalles vanligvis antineoplastisk antibiotika (mot kreft). Actinomycin D, for eksempel, påvirker topoisomerase II og brukes i Wilms -svulster hos barn og i Rabdomiosarcomas.
Antracykliner
Antracykliner er, blant antibiotika, av de mest effektive anti -kreftmedisiner og bredere spekter. De brukes til behandling av lungekreft, eggstokker, livmor, mage, blære, bryst, leukemi og lymfomer. Det er kjent at det påvirker topoisomerase II ved interkalasjon i DNA.
Den første isolerte antracyklinen til en actinobacteria (Streptomyces Peucetius) Det var daunorubicin. Deretter ble doxorubicin syntetisert i laboratoriet, og epirubicine brukes også, og idarubicine.
Antraquinoner
Antraquinoner eller antracenedas er forbindelser avledet fra antracen, ligner antracykliner, som påvirker aktiviteten til topoisomerase II ved interkalasjon i DNA. De brukes til metastatisk brystkreft, ikke -Hodgkinian lymfom (LNH) og leukemi.
Disse medisinene ble funnet i pigmentene til noen insekter, planter (Frogula, Sen, Rhibarb), lav og sopp; så vel som i La Hoelita, som er et naturlig mineral. Avhengig av dosen din, kan de være kreftfremkallende.
Blant disse forbindelsene har vi Mythoxantrona og dets analog Losxantrona. Disse forhindrer spredning av ondartede tumorceller, og blir med DNA irreversibelt.
Epidofilotoksiner
Podofilotoksiner, så som epidofilotoksiner (VP-16) og Teniposide (VM-26), danner et kompleks med topoisomerase II. De brukes mot lunge, testikulær kreft, leukemi, lymfomer, eggstokkreft, brystkarsinom og ondartede intrakranielle svulster, blant andre. Er isolert fra planter Podophyllum notatum og P. Peltatum.
Camptotecinas -analoger
Campotecinas er forbindelser som hemmer topoisomerase I, og blant dem er irinotecan, topoten.
Disse forbindelsene har blitt brukt mot tykktarms-, lunge- og brystkreft, og er naturlig oppnådd fra skorpen og bladene på treslaget Camptotheca Accuminata av kineserne og Tibet Cornacious.
Naturlig hemming
Strukturelle endringer av topoisomeraser I og II kan også forekomme på en helt naturlig måte. Dette kan skje under noen hendelser som påvirker din katalytiske prosess.
Blant disse endringene kan dannelsen av pyrimidin -diametre, misforhold av nitrogenbaser og andre hendelser forårsaket av oksidativt stress siteres.
Referanser
- Liu, l. F. (1994). DNA Topoisomerase: Topoisomerase-målrettingsmedisiner. Akademisk presse. s. 307
- Osheroff, n. og bjorsti, m. (2001). DNA -topoisomerase. Enzymologi og medisiner. Vol. Ii. Menneskelig presse. S. 329.
- « Makrofytter Karakteristikker, habitat, klassifisering og betydning
- 20 oppfinnelser av renessansen som forandret verden »