

Antikodón
- 2397
- 264
- Magnus Sander Berntsen
Hva er et antikodon?
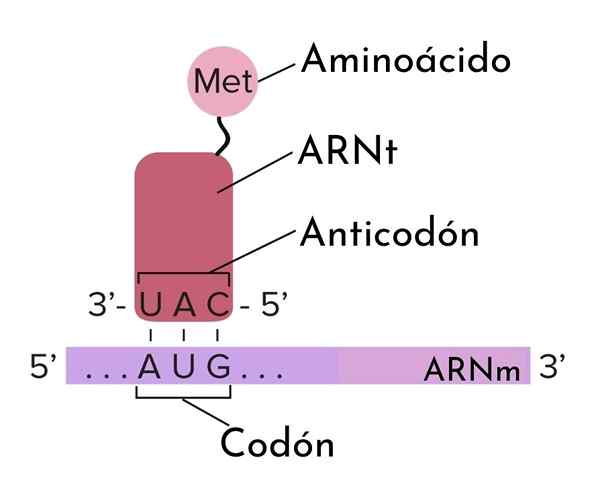
EN Antikodón Det er en sekvens av tre nukleotider som er til stede i et overførings -RNA -moleku.
Denne anerkjennelsen mellom kodoner og antikodoner er antiparallell; Det vil si at den ene er lokalisert i 5 '-> 3' retning mens den andre er festet i en 3 '-> 5' forstand. Denne gjenkjennelsen mellom tre nukleotid (triplett) sekvenser er avgjørende for oversettelsesprosessen; det vil si i syntesen av protein i ribosom.
Under oversettelsen blir messenger -RNA -molekyler således "lest" gjennom anerkjennelsen av kodonene deres av antikodonene til overførings -RNA -ene. Disse molekylene kalles det fordi de overfører en spesifikk aminosyre til proteinmolekylet som dannes i ribosom.
Det er 20 aminosyrer, hver kodet av en spesifikk triplett. Noen aminosyrer er imidlertid kodet av mer enn en triplett.
I tillegg blir noen kodoner gjenkjent av antikodoner i overføring av RNA -molekyler som ikke har noen samlet aminosyre; Dette er de så -kalt stoppkodonene.
Beskrivelse
Et antikodon dannes av en sekvens av tre nukleotider som kan inneholde hvilket som fungerer som en kode.
Antikodoner finnes alltid i overføring RNA-molekyler og er alltid lokalisert i en 3 '-> 5' forstand. Strukturen til disse ARN -ene ligner en kløver, slik at den blir delt inn i fire løkker (eller bånd); I en av løkkene er antikodonet.
Antikodoner er avgjørende for gjenkjennelse av kodonene til messenger -RNA og følgelig for proteinsynteseprosessen i alle levende celler.
Kan tjene deg: multifaktoriell arvAntikodoner fungerer
Hovedfunksjonen til antikodoner er den spesifikke gjenkjennelsen av triplettene som danner kodonene i Messenger RNA -molekyler. Disse kodonene er instruksjonene som er kopiert fra et DNA -molekyl for å diktere rekkefølgen av aminosyrer i et protein.
Når transkripsjon (syntese av kopier av Messenger RNA) forekommer i 5 '-> 3' retning, har kodonene til Messenger RNA denne orienteringen. Derfor må antikodonene som er til stede i overførings-RNA-molekyler ha motsatt orientering, 3 '-> 5'.
Denne foreningen skyldes komplementaritet. For eksempel, hvis et kodon er 5'-AGG-3 ', er antikodonet 3'-UCC-5'. Denne typen spesifikk interaksjon mellom kodoner og antikodoner er et viktig trinn som gjør at nukleotidsekvensen i messenger -RNA kan kode en aminosyresekvens i et protein.
Forskjeller mellom antikodon og kodon
- Antikodoner er trinukleotidenheter i tRNA -er, komplementære til kodonene i mRNA. De lar tRNA levere riktige aminosyrer under proteinproduksjon. I stedet er kodoner trinukleotidenheter i DNA eller RNA, som koder for en spesifikk aminosyre i proteinsyntese.
- Antikodoner er koblingen mellom nukleotidsekvensen til mRNA og aminosyresekvensen til proteinet. Tvert imot overfører kodoner genetisk informasjon fra kjernen der DNA blir funnet til ribosomer der proteinsyntese utføres.
- Antikodonet finnes i antikodonarmen til tRNA -molekylet, i motsetning til kodonene, som er lokalisert i DNA- og RNAM -molekylet.
Kan tjene deg: hemicigososis- Antikodonet er komplementært til det respektive kodonet. På den annen side er kodonet i RNM komplementær til en nukleotidtriplett av et visst gen i DNA.
- Et tRNA inneholder et antikodon. Tvert imot, et mRNA inneholder en rekke kodoner.
Balanseringshypotesen
Balanshypotesen foreslår at fagforeningene mellom det tredje nukleotidet til Messenger RNA -kodonet og det første nukleotidet til overførings -RNA -antikodonet er mindre spesifikke enn leddene mellom de to andre nukleotidene av tripletten.
Crick beskrev dette fenomenet som en "balanse" i den tredje kodonposisjonen. Noe skjer i den posisjonen som gjør at fagforeningene kan være mindre strenge enn normalt. Det er også kjent som Bamboleo eller Tamole.
Denne Cricks Bamboleo -hypotese forklarer hvordan antikodonet til en gitt Arnt kan parre seg med to eller tre forskjellige RNM -kodoner.
Crick foreslo at det å være sammenkoblingen av baser (mellom base 59 av antikodonet i kunst og basen 39 av kodonet i RNM) mindre streng enn normalt, er en viss "bamboleo" eller redusert affinitet tillatt på dette nettstedet.
Som et resultat gjenkjenner en enkelt trin ofte to eller tre av de relaterte kodonene som spesifiserer en gitt aminosyre.
Normalt følger hydrogenbindingene mellom basene til ARNT -antikodonene og RNM -kodonene strenge regler for baseparring bare for de to første basene i kodonet. Imidlertid forekommer denne effekten ikke i alle tredje posisjoner av alle ARNM -kodoner.
RNA og aminosyrer
Basert på bamboleo -hypotesen ble eksistensen av minst to overførings -RNA for hver aminosyre med kodoner som viser fullstendig degenerasjon, som har vist seg å være sant, spådd.
Det kan tjene deg: ufullstendig dominans eller semi -midtinensDenne hypotesen spådde også utseendet til tre overførings -RNA for de seks serinkodonene. Faktisk har tre Arnt blitt karakterisert for serinen:
- Art for Serine 1 (Anticodón AGG) blir med på UCU- og UCC -kodonene.
- Art for Serine 2 (Anticodón Agu) blir med i UCA- og UCG -kodonene.
- Art for Serine 3 (Anticodón UCG) binder seg til AGU- og AGC -kodonene.
Disse spesifisitetene ble bekreftet ved den stimulerte foreningen av rensede aminoacil-arnt trinukleotider, til in vitro ribosomer.
Til slutt inneholder flere overføring RNA. Inosine produseres ved en post -beskrivende modifisering av adenosin.
Cricks Bamboleo -hypotese spådde at når inosinen er til stede i 5 'enden av et antikodon (svingningsposisjonen), ville det bli parret med uracil, cytosin eller adenin i kodonet.
Faktisk binder alanil-arnt-renset inneholdende inosin (I) i posisjon 5 'av antikodonet til ribosomer aktivert med trinukleotider av GCU, GCC eller GCA.
Det samme resultatet er oppnådd med annet renset tRNA med Inosina i posisjon 5 'av antikodonet. Derfor forklarer Cricks Bamboleo -hypotese forholdet mellom ARN og kodoner gitt den genetiske koden, som er degenerert, men bestilt.
Referanser
- Brooker, r. (2012). Konsepter om genetikk (1. utg.). McGraw-Hill Companies, Inc.
- Brown, t. (2006). Genomer 3 (3Rd). Garland Science.
- Griffiths, a., Wessler, s., Carroll, s. & Doebley, j. (2015). Introduksjon til genetisk analyse (11. utg.). W.H. Freeman
- Lewis, r. (2015). Menneskelig genetikk: konsepter og applikasjoner(11. utg.). McGraw-Hill Education.
- Snustad, d. & Simmons, m. (2011). Prinsipper for genetikk(6. utg.). John Wiley og sønner.