

Beta oksidasjon av fettsyrer trinn, reaksjoner, produkter, regulering
- 4010
- 711
- Dr. Andreas Hopland
De Beta oksidasjon av fettsyrer Det er ruten for katabolisme (nedbrytning) av fettsyrer, som har som hovedfunksjon produksjonen eller "frigjøringen" av energien som finnes i koblingene til disse molekylene.
Denne ruten ble oppdaget i 1904 takket være eksperimentene utført av den tyske Franz -knoopen, som besto av administrasjonen, eksperimentelle rotter, av fettsyrer hvis endelige metylgruppe hadde blitt modifisert med en fenylgruppe.
Beta skjema oksidasjon av fettsyrer (kilde: Arturo González Laguna [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Knoop forventet katabolismeproduktene fra disse "analoge" fettsyrene til å følge ruter som ligner på oksidasjonsveien til normale fettsyrer (de umodifiserte innfødte). Han fant imidlertid at det var forskjeller i produktene som ble oppnådd avhengig av antall karbonatomer av fettsyrer.
Med disse resultatene foreslo Knoop at nedbrytning skjedde i "trinn", og startet med et "angrep" β -karbon (posisjon 3 med hensyn til terminalkarboksylgruppen), og frigjorde fragmenter av to karbonatomer.
Deretter ble det vist at prosessen krever energi i form av ATP, som oppstår i mitokondrier og at fragmentene av to karbonatomer kommer inn i Krebs-syklusen som acetyl-CoA.
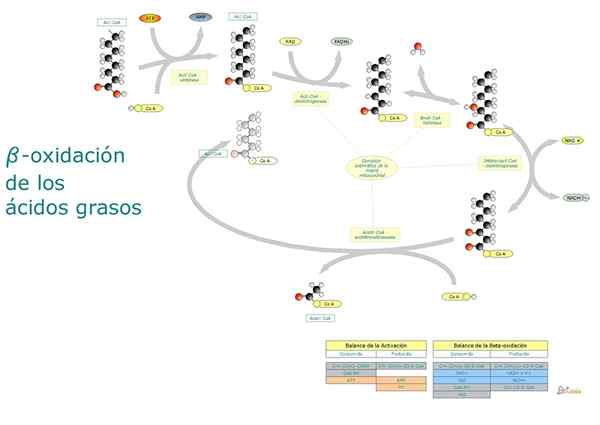
Kort sagt, beta -oksidasjon av fettsyrer innebærer aktivering av den terminale karboksylgruppen, transport av fettsyre aktivert mot mitokondriell matrise og den "forskjøvede" oksidasjonen av to i to karbonatomer fra karboksylgruppen.
Som mange anabole og katabolske prosesser er denne ruten regulert, siden den fortjener mobilisering av fettsyrer av "reserve" når de andre katabolske rutene ikke er nok til å møte celle- og kroppsenergibehov.
[TOC]
Trinn og reaksjoner
Fettsyrer er hovedsakelig i cytosol, de kommer allerede fra biosyntetiske ruter eller fettforekomster som er lagret fra inntatt mat (som må komme inn i celler).
- Aktivering av fettsyrer og transport til mitokondrier
Aktivering av fettsyrer krever bruk av et ATP -molekyl og har å gjøre med dannelsen av acyl -tioés konjugater med koenzym til.
Det kan tjene deg: viktigheten av biologiDenne aktiveringen blir katalysert av en gruppe enzymer kalt acetyl-CoA-spesifikk lengde med hensyn til lengden på kjeden til hver fett. Noen av disse enzymene aktiverer fettsyrer når de blir transportert til mitokondriell matrise, da de er innebygd i den ytre mitokondrielle membranen.
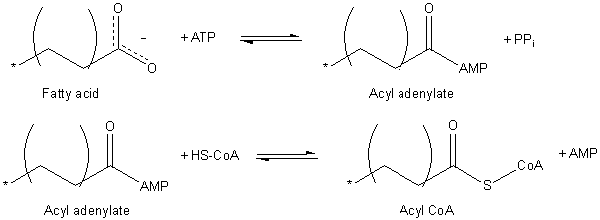 Aktivering av fettsyrer (kilde: JAG123 på engelsk Wikipedia [Public Domain] via Wikimedia Commons)
Aktivering av fettsyrer (kilde: JAG123 på engelsk Wikipedia [Public Domain] via Wikimedia Commons) Aktiveringsprosessen skjer i to trinn, og produserer først en adenylat acyl fra ATP -fettsyren, hvor et pyrofosfatmolekyl (PPI) frigjøres. Karboksylgruppen aktivert av ATP blir senere angrepet av TIOL-gruppen av koenzymet ved å danne ACIL-CoA.
ACIL-CoA-translokasjon gjennom mitokondriemembranen oppnås takket være et transportsystem kjent som Carnitine Shuttle.
- Beta oksidasjon av mettede fettsyrer med et par karbonatomer
Nedbrytningen av fettsyrer er en syklisk rute, siden frigjøringen av hvert fragment av to karbonatomer umiddelbart blir fulgt av en annen, inntil den totale lengden på molekylet. Reaksjonene som har del i denne prosessen er følgende:
- Dehydrogenering.
- Hydrering av en dobbeltbinding.
- Dehydrogenering av en hydroksylgruppe.
- Fragmentering for angrepet av et acetyl-CoA-molekyl på karbon β.
Reaksjon 1: Første dehydrogenering
Den består av dannelsen av en dobbeltbinding mellom α -karbonet og ß -karbonet ved eliminering av to hydrogenatomer. Det katalyseres av et acil-CoA dehydrogenase-enzym, som danner et trans-trans-molekyl transmolekyl.
Reaksjoner 2 og 3: Hydrering og dehydrogenering
Hydrering katalyseres av engelbelegget+.
Hydrering av transhvite.
FADH2 og NADH produserte i de tre første reaksjonene av beta -oksidasjonen blir gjenvist.
Det kan tjene deg: Ovogonias: Hva er, beskrivelse, morfologiReaksjon 4: Fragmentering
Hver oksidasjonsbetasyklus som eliminerer et molekyl av to karbonatomer ender.
Denne reaksjonen katalyseres av ß-cototoolase eller tisolase-enzymet, og produktene er et acil-CoA-molekyl (fettsyren aktivert med to mindre karbonatomer) og en av acetyl-CoA.
- Beta oksidasjon av mettede fettsyrer med et rart antall karbonatomer
I de fettsyrene i det rare antallet karbonatomer (som ikke er veldig rikelig), har molekylet i den siste nedbrytningssyklusen 5 karbonatomer, så fragmenteringen produserer et acetyl-CoA-molekyl (som kommer inn i syklusen av Krebs) og en annen av Propionil-CoA.
Propionyl-CoA må være karboksylert (ATP og bikarbonatavhengig reaksjon) ved propionyl-CoA karboksylaseenzym, som danner en forbindelse kjent som D-metylmalonyl-CoA, som må epimeriseres til sin form "L".
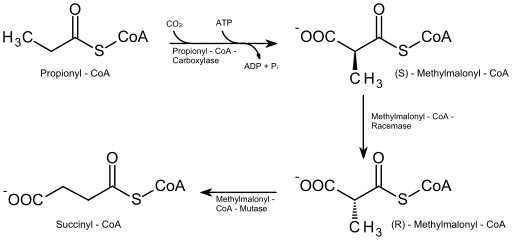 Betaoksidasjon av fettsyrer av oddetall (kilde: Eleska [CC0] via Wikimedia Commons)
Betaoksidasjon av fettsyrer av oddetall (kilde: Eleska [CC0] via Wikimedia Commons) Forbindelsen som følge av epimeriseringen blir senere omdannet til succinyl-CoA ved virkning av enzymet L-metylmalonyl-CoA-mutase, og dette molekylet, så vel som acetyl-CoA, kommer inn i syklusen av sitrussyrer.
- Beta oksidasjon av umettede fettsyrer
Mange cellelipider har fettsyrekjeder med umettethet, det vil si at de har en eller flere dobbeltbindinger mellom karbonatomene.
Oksidasjonen av disse fettsyrene er litt forskjellig fra mettede fettsyrer, ettersom to ekstra enzymer, et isomerase sinne og 2,4-dieno-CoA-reduktase, er ansvarlige for Enzymet-CoA hydratasa.
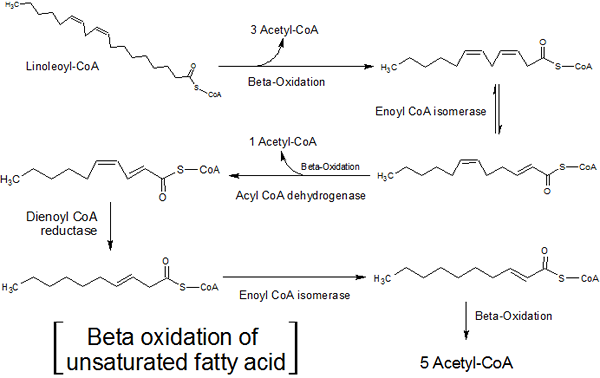 Betaoksidasjon av umettede fettsyrer (kilde: Hajime7basketball [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons)
Betaoksidasjon av umettede fettsyrer (kilde: Hajime7basketball [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) Isomerase-sinne virker på enumettede fettsyrer (med en enkelt umettethet), i mellomtiden reagerer enzymet 2,4-dieno-CoA-reduktase med flerumettede fettsyrer (med to eller flere umettet)).
- Beta ekstra -bane oksidasjon
Beta oksidasjon av fettsyrer kan også forekomme i andre cytosoliske organeller som peroksisomer, for eksempel med forskjellen at elektroner som overføres til FAD+ ikke blir levert til luftveiene, men direkte til oksygen.
Kan tjene deg: Alstroemeria: Kjennetegn, habitat, omsorg, arterDenne reaksjonen produserer hydrogenperoksyd (oksygen reduseres), forbindelse som elimineres av katlaseenzymet, spesifikt for disse organellene.
Beta oksidasjonsprodukter
Oksidasjonen av fettsyrer gir mye mer energi enn karbohydratforringelse. Hovedproduktet av beta-oksidasjon er acetyl-CoA produsert i hvert trinn i den sykliske delen av ruten, men andre produkter er imidlertid:
- AMP, H+ og pyrofosfat (PPI), produsert under aktivering.
- FADH2 og NADH, for hver produsert acetyl-CoA.
- Succinil-CoA, ADP, PI, for rare kjedesyrer.
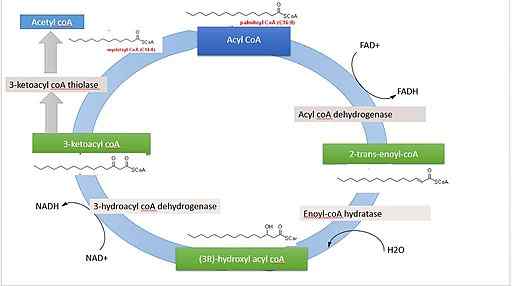 Betaoksidasjon av palmitinsyre (kilde: 'Rojinbkht [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Betaoksidasjon av palmitinsyre (kilde: 'Rojinbkht [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Hvis vi vurderer fullstendig oksidasjon av palmitinsyre (palmitat) som et eksempel, er en fettsyre med 16 karbonatomer, mengden energi som oppstår mer eller mindre tilsvarer 129 ATP -molekyler, som kommer fra de 7 omgangene, må fullføre den syklus.
Regulering
Regulering av beta -oksidasjon av fettsyrer i de fleste celler avhenger av energitilgjengelighet, ikke bare relatert til karbohydrater, men med de samme fettsyrene.
Dyr kontrollerer mobilisering og derfor nedbrytning av fett av hormonelle stimuli, som samtidig kontrolleres av molekyler som AMPC, for eksempel.
I leveren, det viktigste fettnedbrytningsorganet, er konsentrasjonen av malonyl-CoA ekstremt viktig for regulering av beta-oksidasjon; Dette er det første underlaget som er forpliktet til biosyntese -ruten til fettsyrer.
Når malonyl-CoA samler seg i store proporsjoner, fremmer det fettsyrebiosyntese og hemmer mitokondriell transportør eller acyl-karnitin-skyttelbuss. Når konsentrasjonen avtar, opphører hemming og beta -oksidasjonen aktiveres.
Referanser
- Mathews, c., Van Holde, K., & Ahern, K. (2000). Biokjemi (3. utg.). San Francisco, California: Pearson.
- Nelson, d. L., & Cox, m. M. (2009). Lehninger prinsipper for biokjemi. Omega -utgaver (5. utg.).
- Rawn, J. D. (1998). Biokjemi. Burlington, Massachusetts: Neil Patterson Publisher.
- Schulz, h. (1991). Beta oksidasjon av fettsyrer. Biochimica et Biophysica Acta, 1081, 109-120.
- Schulz, h. (1994). Regulering av fettsyreoksidasjon i hjertet. Kritisk anmeldelse, 165-171.
- Schulz, h., & Kunau, w. (1987). Beta-oksidasjon av umettede fettsyrer: en revice-bane. Tibs, 403-406.