

Kalsiumpumpefunksjoner, typer, struktur og drift
- 3746
- 906
- Jonathan Moe
De Kalsiumpumpe Det er en protein naturstruktur som er ansvarlig for transport av kalsium gjennom cellemembraner. Denne strukturen er ATP -avhengig og regnes som et ATPASA -type protein, også kalt CA2+-Atpasa.
Ca2+-Atpasa finnes i alle celler i eukaryote organismer og er avgjørende for kalsiumhomeostase i cellen. Dette proteinet utfører primær aktiv transport, siden bevegelsen av kalsiummolekyler går mot konsentrasjonsgradienten.
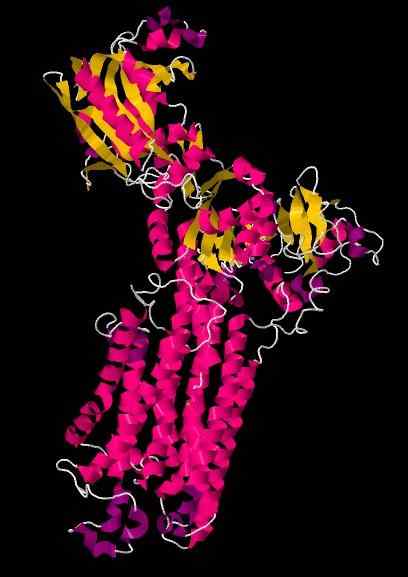
Serca krystallografisk struktur.
Kilde: WCNSAFFO [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] [TOC]
Kalsiumpumpefunksjoner
Ca2+ Det oppfyller viktige artikler i cellen, så deres regulering i dem er grunnleggende for deres riktige drift. Fungerer ofte som en annen messenger.
I ekstracellulære rom konsentrasjonen av Ca2+ Det er omtrent 10.000 ganger høyere enn innenfor celler. En økning i konsentrasjonen av dette ionet i cellcytoplasma utløser forskjellige responser, for eksempel muskelsammentrekninger, frigjøring av nevrotransmitter og nedbrytning av glykogen.
Det er flere måter å flytte disse ionene fra cellene: passiv transport (ikke -spesifikk utgang), ionekanaler (bevegelse til fordel for dens elektrokjemiske gradient), antiportert sekundær aktiv transport (NA/CA) og primær aktiv transport med pumpe -ATP -avhengig.
I motsetning til de andre mekanismene for forskyvning av CA2+, Pumpen fungerer i vektorform. Det vil si at ionen beveger seg i en retning slik at det bare fungerer ved å utvise dem.
Cellen er ekstremt følsom for Ca -konsentrasjonsendringer2+. Ved å presentere en så markant forskjell med sin ekstracellulære konsentrasjon, så er det så viktig å effektivt gjenopprette dets normale cytosoliske nivå.
Kan tjene deg: mesosomFolkens
Tre typer CA er beskrevet2+-Atasas i dyreceller, i henhold til deres lokasjoner i cellene; Pumpene som ligger i plasmamembranen (PMCA), de som ligger i endoplasmatisk retikulum og atommembranen (SERCA), og de som finnes i Golgi -apparatmembranen (SPCA).
SPCA -bomber transporterer også Mn -ioner2+ som er kofaktorer av forskjellige enzymer av matrisen til Golgi -apparatet.
Gjærceller, fra andre eukaryoterende organismer og planteceller har andre typer CA2+-Veldig spesielle atasas.
Struktur
PMCA -pumpe
I plasmamembranen finner vi den aktive antiportitransporten Na/Ca, og er ansvarlig for forskyvningen av en betydelig mengde CA2+ I hvile- og aktivitetsceller. I de fleste celler i ro er manageren for transport av kalsium utover PMCA -pumpen.
Disse proteinene består av omtrent 1.200 aminosyrer, og har 10 transmembranale segmenter. I cytosol er det 4 hovedenheter. Den første enheten inneholder den aminoterminale gruppen. Det andre har grunnleggende egenskaper, som lar fosfolipider bli med aktivatorer.
I den tredje enheten er det en asparaginsyre med katalytisk funksjon, og "nedstrøms" av dette et fluorescein iso -sykling bindende bånd, i ATP Union -domenet.
I den fjerde enheten er mestring av union til calmodulin, gjenkjennelsesstedene til visse kinaser (A og C) og de bindende båndene til CA2+ Alosterisk.
SERCA PUMP
SERCA -pumper er i stor mengde i sarkoplasmatisk retikulum av muskelceller, og deres aktivitet er relatert til sammentrekning og avslapning i muskelbevegelsessyklusen. Funksjonen er å transportere CA2+ Fra cytosolen til cellen til retikulummatrisen.
Kan tjene deg: Basal Stratum: Karakteristikker og funksjonerDisse proteinene består av en enkelt polypeptidkjede med 10 transmarkedsdomener. Strukturen er i utgangspunktet lik den for PMCA -proteiner, men den skiller seg ut ved at de bare har tre enheter i cytoplasmaet, og er det aktive stedet i den tredje enheten.
Driften av dette proteinet krever en belastningsbalanse under ionetransport. To ca2+ (av ATP hydrolysert) forskyves fra cytosol til retikkelmatrisen, mot en veldig høy konsentrasjonsgradient.
Denne transporten skjer antiportisk, for samtidig to H+ De blir rettet mot cytosol fra matrisen.
Operasjonsmekanisme
Serca -bomber
Transportmekanismen er delt inn i to E1- og E2 -stater. I E1 -fagene som presenterer en høy affinitet for CA2+ De er rettet mot cytosol. I E2 er unionsstedene rettet mot lumen i retikulum som presenterer en lav affinitet for CA2+. De to CA -ionene2+ Kom sammen etter overføringen.
Under unionen og overføringen av CA2+, Konformasjonsendringer skjer, inkludert åpningen av proteinets m -domene, som er mot cytosol. Ionene blir deretter lettere bundet til de to unionsstedene til nevnte domene.
Unionen av de to CA -ionene2+ fremmer en serie strukturelle endringer i protein. Blant dem rotasjonen av visse domener (domene a) som omorganiserer bombeenhetene, slik at åpningen mot matrisen til retikulum skal frigjøre ionene, som er frakoblet takket være reduksjonen i affinitet i unionsstedene.
Protonene h+ og vannmolekyler stabiliserer unionsstedet til CA2+, som får domenet til å rotere igjen til sin opprinnelige tilstand, og lukker tilgangen til endoplasmatisk retikulum.
Kan tjene deg: cariocinesisPMCA -pumper
Disse typer pumper finnes i alle eukaryote celler og er ansvarlig for utvisning av CA2+ mot det ekstracellulære rommet for å holde konsentrasjonen i cellene stabil.
I dette proteinet transporteres et ca ion2+ Av hydrolysert ATP. Transport reguleres av nivåene av calmodulinproteinet i cytoplasma.
Ved å øke CA -konsentrasjonen2+ Cytosolisk, calmodulinnivået øker, som er bindende til kalsiumioner. CA -komplekset2+-Calmodulin, samler deretter PMCA Bomb Bomb Site. Det er en konformasjonsendring i pumpen som gjør at åpningen til det ekstracellulære rommet kan utsettes.
Kalsiumioner frigjøres, og gjenoppretter normale nivåer inne i cellen. Følgelig CA -komplekset2+-Calmodulin er desasambla, og returnerer pumpen til sin opprinnelige tilstand.
Referanser
- Brini, m., & Carafoli, og. (2009). Kalsiumpumper i helse og sykdom. Fysiologiske anmeldelser, 89(4), 1341-1378.
- Carafoli, e., & Brini, m. (2000). Kalsiumpumper: Strukturelt grunnlag for og mekanisme for kalsiumtransmembrantransport. Nåværende mening i kjemisk biologi, 4(2), 152-161.
- Devlin, t. M. (1992). Biokjemiets lærebok: med kliniske korrelasjoner.
- Latorre, R. (Red.). (nitten nittiseks). Biofysikk og cellefysiologi. Sevilla University.
- Lodish, h., Darnell, J. OG., Berk, a., Kaiser, ca. TIL., Krieger, m., Scott, m. P., & Matsudaira, P. (2008). Mollekylær cellebiologi. Macmillan.
- Pocock, g., & Richards, C. D. (2005). Menneskelig fysiologi: Medisinsk base. Elsevier Spania.
- Voet, d., & Voet, j. G. (2006). Biokjemi. Ed. Pan -American Medical.
- « Eumetazoa -egenskaper, taksonomi og klassifisering
- Colombianske furuegenskaper, habitat, reproduksjon og bruk »