

Elektron transportørkjedekomponent, sekvens, hemmere
- 2473
- 697
- Mathias Aas
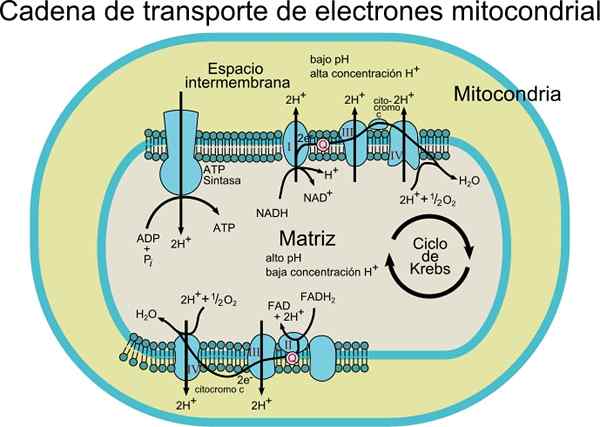
De Elektron transportørkjede Det består av et sett med protein- og koenzymmolekyler inne i en membran. Som navnet tilsier, er det ansvarlig for transport av elektroner fra NADH- eller FADH2 -koenzymer til den endelige mottakeren som er O2 (molekylært oksygen).
I denne transportprosessen er energien som frigjøres ved å overføre elektroner fra koenzymer til molekylært oksygen av redoksentre United til protein, assosiert med energiproduksjon (ATP). Denne energien oppnås takket være protongradienten som genereres i den mitokondrielle indre membranen.
Dette transportsystemet består av forskjellige komponenter som finnes i minst to oksidasjonstilstander. Hver av dem er effektivt redusert og reoxyD under bevegelsen av elektroner fra NADH eller FADH2 til O2.
NAD+ og FAD -koenzymer reduseres i fettsyreoksidasjonsveier og sitronsyresyklusen som et resultat av oksidasjon av forskjellige underlag. Deretter oksideres disse koenzymene i den elektroniske transportkjeden.
Slik at det elektroniske transportsystemet består av en sekvens av oksydreduksjonsreaksjoner som er koblet til hverandre.
[TOC]
Kjedekomponenter
Avhengig av typen organisme, kan 3 til 6 komponenter observeres som utgjør elektrontransportørkjeden. Elektrontransportprosessen og ATP -syntese ved oksidativ fosforylering er prosesser som oppstår i en membran.
Når det gjelder prokaryote celler (aerobe bakterier), skjer disse prosessene assosiert med plasmamembranen. I eukaryotceller forekommer det i mitokondriell membran, så elektrontransportkomponentene finnes i den indre delen av membranen.
https: // giphy.COM/GIFS/HTKSBBEDQUK9Z7CO5E
Elektroner overføres gradvis gjennom fire komplekser som utgjør den elektroniske transportkjeden.
Hvert kompleks har flere proteinkomponenter assosiert med protesegrupper (ikke -aminosyrekomponenter av konjugatproteiner) redoks, som lar reduksjonspotensialene øke.
I tillegg består dette transportsystemet av forskjellige molekylære arter som flavoprotein; koenzym som også kalte Ubiquinona (COQ eller UQ); forskjellige cytokromer som cytokrom B, C, C1, A og A3; Proteiner med Fe-S og proteingrupper knyttet til Cu. Disse molekylene er knyttet til membranen, med unntak av cytokrom c.
Kompleks i
https: // giphy.com/gifs/rlf1rl9gdc6356sj6w
I-komplekset kalte NADH Coenzyme Quinona Oxidortaduase, eller NADH dehydrogenase, består av rundt 45 polypeptidkjeder og inneholder et mononukleotid flavinmolekyl (FMN) og åtte til ni Fe-S-grupper Fe-S-S. Som navnet tilsier, overfører dette komplekset et par elektroner fra NADH -koenzymet til COQ.
Funksjonen til NADH dehydrogenase -komplekset begynner med foreningen av NADH til nevnte kompleks på siden av den indre mitokondrielle membranmatrisen. Elektronene blir deretter transportert fra NADH til FMN. Deretter passerer elektroner fra det reduserte flavinet (FMNH2) til Fe-S-proteiner.
FMNH2 fungerer som en slags bro mellom NADH og FE-S-proteinene, siden sistnevnte bare kan overføre et enkelt elektron, mens NADH-koenzymet overfører to, slik at Flavinas gjør denne overføringen av et enkelt elektron takket være dets redoks av semi -kynonen.
Kan tjene deg: monocytopoiesis: stadier, egenskaper, reguleringEndelig blir elektronene overført fra FE-S-grupperingene til koenzymet Q, som er en mobil elektrontransportør med en isoprenidhale som gjør den hydrofob som tillater å krysse midten av den mitokondrielle membranen.
Kompleks II
![]()
II -komplekset, bedre kjent som succinatdehydrogenase, er et integrert protein av den mitokondrielle indre membranen, og er et enzym som griper inn i sitronsyresyklus.
Dette komplekset består av to hydrofile underenheter og to hydrofobe med hemo B-grupper som gir COQ-bindingsstedet, i tillegg til et flavoprotein og et protein med Fe-S.
I sitronsyresyklusen (Krebs eller Tricarboxylic Acid Cycle) omdannes succinat til fumarat ved succinatdehydrogenase, noe som reduserer koenzymfadet til FADH2. Fra dette siste koenzymet overføres elektroner til FE-S-sentrene som igjen overfører dem til COQ.
Under reaksjonene fra denne elektronoverføringen er standard redokspotensialet veldig lavt, noe som forhindrer at den nødvendige frie energien blir frigjort for å syntetisere ATP.
Dette betyr at Complex II er det eneste komplekset i elektrontransportkjeden som ikke kan gi energi til ATP -syntese. Imidlertid er dette komplekset nøkkelen i prosessen, siden det overfører elektroner fra FADH2 til resten av kjeden.
Kompleks III
https: // giphy.com/gifs/cjg0raiuizjds2hkyu
III -komplekset, cytokromkompleks BC1 eller CoQ cytokrom c -reduktase, overfører elektronene fra det reduserte koenzym til cytokrom c. Denne overføringen skjer gjennom en enkelt redoksvei, som er kjent som Q -syklusen.
Dette komplekset består av et protein med Fe-S og tre forskjellige cytokromer, der jernatomet som ligger i hemo-gruppen varierer syklisk mellom de reduserte tilstandene (Fe2+) og oksidert (Fe3+).
Cytokromer er hemoproteiner av elektronisk transport, som har redoksaktivitet. De er til stede i alle organismer, med unntak av noen obligatoriske anaerober.
Disse proteinene har hemo -grupper som veksler mellom to oksidasjonstilstander (Fe2+ og Fe3+). Cytochrome C er en mobil elektrontransportør som er svakt assosiert med den indre membranen til mitokondrier.
Cytokromene som er funnet i dette komplekset er cytokromer B, C og A, de 3 er aktive redoksproteiner med grupper vi har forskjellige egenskaper, som veksler oksidasjonstilstandene deres mellom Fe2+ og Fe3+.
Cytokrom C er et perifer membranprotein som fungerer som en "skyttelbuss" av elektron med cytokrom C1 og med IV -komplekset.
IV -kompleks
Cytokrom C og O2 er de endelige reseptorene av elektroner avledet fra oksidasjon av organisk materiale, så IV- eller cytokrom C -oksidasekomplekset er det terminale enzymet i den elektroniske transportprosessen. Dette aksepterer elektroner fra cytokrom c og overfører dem til O2 -reduksjon.
Den komplekse funksjonen er å katalysere oksidasjoner av et elektron av de fire påfølgende molekylene av redusert cytokrom C, det vil si samtidig reduserer fire elektroner av et O2 -molekyl, og til slutt produserer to H2O -molekyler.
Kan tjene deg: epitelbeleggvevElektrontransportsekvens
Elektronene blir overført fra kompleksene I og II til komplekset III takket være koenzymet Q, og derfra passerer de til IV -komplekset gjennom cytokrom c. Når elektronene går gjennom disse fire kompleksene, øker reduksjonspotensialet, og frigjør energi, som deretter brukes til ATP -syntese.
Totalt forårsaker overføringen av et par elektroner translokasjon av 10 protoner gjennom membranen; fire i komplekser I og IV og to av komplekset III.
Nadh Descidrogenasa
Dette enzymet katalyserer oksidasjonen av NADH -koenzym av koenzymet som. Elektronene beveger seg fra NADH til FMN som er festet til den hydrofile halen til komplekset I. FE-S-grupperinger overfører til elektroner om gangen. Disse FE-S-gruppene reduserer til COQ, som er innebygd i membranen, til Ubiquinol (redusert COQ).
Under overføringen av elektroner til COQ overføres fire protoner igjen gjennom den indre membranen, til det intermembranale rommet. Mekanismen som disse protonene er translokert innebærer protein lokalisert i den hydrofobe halen til komplekset I.
Overføringsprosessen for elektroner i dette trinnet frigjør gratis energi, spesielt -16,6 kcal/mol.
CoQ-Citocromo C-reduktase og Q-syklusen
Koenzym som oksideres av cytokrom C, i en reaksjon katalysert av dette koenzym. Oksidasjonen av ubiquinol (redusert COQ) forekommer på et bestemt sted av komplekset (QO- eller oksidasjonssted) i mitokondriell membran, og overfører to elektroner, en til proteinet med Fe-S-gruppene og en annen til hemokruppene.
I Q -syklusen produserer oksidasjonen av COQ semi -quinon, og det er her elektronene blir overført til hemo B1- og BH -gruppene. Mens denne elektronoverføringen oppstår, oksiderer en andre COQ på QO -stedet, og gjentar syklusen.
Denne syklusen forårsaker overføring av to elektroner og på sin side translokasjon av fire protoner til det intermembranale rommet, med frigjøring av -10,64 kcal/mol gratis energi.
Citokrom C -oksidase
Dette enzymet (kompleks IV) katalyserer oksidasjonen av cytokrom c (redusert) med O2, som er den endelige elektronen akseptor. Denne overføringen produserer et H2O -molekyl for hvert par elektroner som overføres i tillegg til translokasjon av protoner gjennom membranen.
Elektronene beveger seg fra en til en, fra den reduserte cytokrom C til et par Cua -ioner, deretter passerer de til en hemo en gruppe og til slutt når binuclear sentrum av komplekset som inneholder Cub og Hemo A3 -ioner, hvor overføringen av fire Elektroner oppstår for oksygen.
I IV -komplekset overfører elementene elektronene til en etter en, slik at O2 reduseres gradvis, slik at frigjøring av noen giftige forbindelser som superoksyd, hydrogenperoksyd eller hydroksylradikaler ikke forekommer.
Energien som frigjøres på dette stadiet tilsvarer -32 kcal/mol. Den elektrokjemiske gradienten som ble generert under overføringsprosessen og energiforandringene (ΔE) forårsaket av et par elektroner når de passerer gjennom de fire kompleksene, tilsvarer på hvert trinn til den frie energien som er nødvendig for produksjon av et ATP -molekyl.
Kan tjene deg: membranfoldteoriStor dehydrogenase
Som nevnt har dette komplekset den eneste, men viktige funksjonen å introdusere FADH2 -elektronene fra sitronsyresyklusen til elektrontransportkjeden.
Dette enzymet katalyserer oksidasjonen av FADH2 -koenzym av koenzymet Q (oksidert). I sitronsyresyklusen når succinatet til fumarat, blir to elektroner og to protoner overført til kjepphest. Deretter overfører FADH2 disse elektronene til COQ gjennom FE-S-sentrene i komplekset.
Til slutt, fra COQ, blir elektronene overført til det komplekse III, etter trinnene beskrevet ovenfor.
Kjedekompleksene er uavhengige
De fire kompleksene som utgjør den elektroniske transportkjeden er uavhengige, det vil si at de er uavhengig og opererer i den mitokondriale indre membranen, og bevegelsen til hver av dem i membranen er ikke avhengig eller er knyttet til de andre kompleksene.
I- og II -kompleksene beveger seg i membranen ved å overføre elektronene sine til COQ som også sprer seg i membranen og overfører dem til komplekse III, hvor elektroner passerer til cytokrom c som også er mobil i membranen og avsetter elektroner i IV -komplekset.
Elektroniske transportkjedehemmere
På den elektroniske transportkjeden virker noen spesifikke hemmere i prosessen. Rothenone er et ofte brukt insektmiddel som er støkiometrisk forbundet med kompleks I, og unngår reduksjon av COQ.
Noen barbiturater type medisiner som piericidine og amytal, hemmer komplekset I, og forstyrrer overføringen av elektroner fra FE-S-gruppene til COQ.
I kompleks II fungerer noen forbindelser som TheoilTrifluoroacetone og Malonate som konkurrerende hemmere med succinat, og unngår deres oksidasjon og i sin tur overføring av elektroner til FAD.
Noen antibiotika som myxotiazol og stigmatellin, binder seg til Q-stedet for COQ, og hemmer overføringen av elektroner fra koenzymet Q til Fe-S-sentrene til proteinene.
Cyanid, azida (N3-), svovelsyre og karbonmonoksid hemmer IV-komplekset. Disse forbindelsene binder seg til hemokrupper, og unngår overføring av elektroner til det komplekse eller oksygen (O2) binuclear Center (O2).
Når elektrontransportørkjeden blir hemmet, stoppes energiproduksjonen på grunn av oksidativ fosforylering, noe som forårsaker alvorlig skade og til og med organismenes død.
Referanser
- Alberts, f., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, P. (2004). Essensiell cellebiologi. New York: Garland Science. 2. utgave.
- Cooper, g. M., Hausman, r. OG. & Wright, n. (2010). Cellen. (s. 397-402). Ed. Marbán.
- Devlin, t. M. (1992). Biokjemiets lærebok: med kliniske korrelasjoner. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, C. M. (2008). Biokjemi. Ed. Thomson Brooks/Cole.
- Rawn, J. D. (1989). Biokjemi (nei. 577.1 rå). Ed. Interamerikansk-McGraw-Hill
- Voet, d., & Voet, j. G. (2006). Biokjemi. Ed. Pan -American Medical.
- « Elektriske Anguila -egenskaper, habitat, reproduksjon
- Biozoosegenskaper, morfologi, reproduksjon, ernæring »