

Calmodulin Structure, Functions and Mechanism of Action
- 2730
- 662
- Markus Fredriksen
Calmodulin Det er et begrep som betyr "kalsiummodulert protein", og refererer til et lite intracellulært protein som eier egenskapen til kalsiumion (Ca ++) og å formidle mange av dets intracellulære handlinger. Opprinnelsen til ordet er født fra kombinasjonen av engelske ord 'kalsium', 'modulert' og 'protein' som, når sammen, stammer i KalkCium ModulAted proteI.
Blant mineralelementene som kommer til å være en del av konstitusjonen av dyreorganismer, er kalsium, etterfulgt av fosfor, mye forskjellig fra det mest rikelig, siden beinet er dannet av avsetningen i sin matrise av store mengder mineralsalter dannet av dette ion.
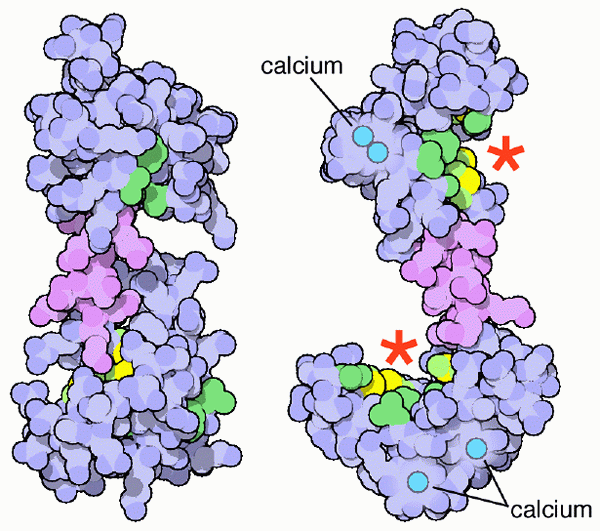
Calmodulin Scheme og dets kalsiumforeningssteder (Kilde: PDB [CC BY-SA (https: // CreativeCommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Selvfølgelig er disse kalsiummineralsaltene avgjørende for grunnloven og konformasjonen av skjelettsystemet til virveldyr, men det er den ioniserte formen for kalsium (Ca ++) i løsning i kroppsvæsker som blir en fysiologisk betydning som er relevant for livet til den Livet til organismer.
Denne kationen, med to positive elektriske ladninger i sin struktur, kan fungere som en nåværende transportør når du beveger seg gjennom cellemembranen og modifiserer nivået av elektrisk potensial i mange av kroppens spennende celler, hovedsakelig i hjertemuskelen.
Men av større fysiologisk relevans er det faktum at mange cellulære regulatoriske reaksjoner avfyrt av ytre stimuli, som nevrotransmittere, hormoner eller andre fysiske eller biokjemiske faktorer, er noen av metabolske fossefall der flere proteiner sekvensielt deltar, noen av dem er enzym som er enzymene som er enzymer som er enzymer, trenger de kalsium for aktivering eller inaktivering.
Det sies da i disse tilfellene at kalsium fungerer som en annen messenger i et metabolsk fossefall som tar sikte på å ha et endelig resultat som ville bli den nødvendige celleresponsen for å imøtekomme et behov som ble oppdaget på et annet nivå annet enn selve cellen, og som krever av henne det spesielle svaret.
Kalsium kan virke direkte på sitt biokjemiske mål for å påvirke aktiviteten, men det krever ofte deltakelse av et protein som det må forenes for å kunne utøve sin effekt på proteinet (e) for å modifisere. Calmodulin er et av de medierende proteiner.
[TOC]
Struktur
Calmodulin, av en stor allestedsnærværende, siden det kommer til uttrykk i nesten alle celletyper av eukaryote organismer, er det et lite syreprotein med omtrent 17 kDa molekylvekt, hvis struktur er ekstremt bevart blant arten.
Kan tjene deg: ioniske kanaler: struktur, funksjoner, typerDet er et monomert protein, det vil si at det dannes av en enkelt polypeptidkjede, som vedtar ved sin terminal ender formen til kuleformet domener forenet av hverandre av en alfa -propell. Hvert kuleformet domene har to grunner kjent som EF -hånd (fra engelsk Ef Hånd) som er typisk for proteinene som binder seg til kalsium.
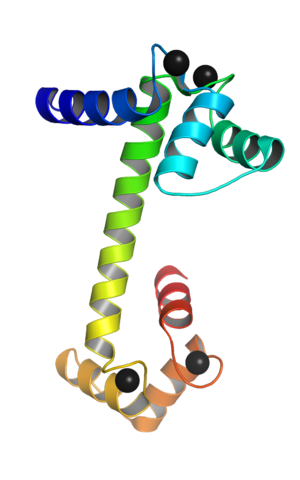 Calmodulin koblet til kalsiumioner (Kilde: Webridge [CC By-SA (https: // CreativeCommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Calmodulin koblet til kalsiumioner (Kilde: Webridge [CC By-SA (https: // CreativeCommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Disse "EF" topologiske motivene representerer en slags supersekundariestrukturer; De er knyttet til hverandre, i hvert kuleformet domene, av et område med stor fleksibilitet, og i hver av dem er det et veikryss for Ca ++, som kaster 4 steder totalt for hvert calmodulinmolekyl.
Foreningen av kalsiumioner, med positiv belastning, er mulig takket være tilstedeværelsen av aminosyreavfall med sidekjeder som er negativt ladet på kalsiumforeningsstedene til calmodulin -kalsium. Dette avfallet er tre aspartater og et glutamat.
Calmodulin -funksjoner
Alle funksjonene så langt kjent for calmodulin er innrammet i en gruppe handlinger fremmet av økningen av cytosolisk kalsium produsert ved inngangen fra det ekstracellulære rommet eller dens avkjørsel fra de intracellulære avsetningene: mitokondrier og endoplasmatisk retikulum.
Mange av kalsiumhandlingene blir oppfylt av dette ionet som virker direkte på dets hvite proteiner, som kan være av mangfoldig karakter og funksjoner. Noen av disse proteinene kan ikke påvirkes direkte, men krever at kalsium blir med på calmodulin, og det er dette komplekset som virker på proteinet påvirket av ionet.
Det sies om disse hvite proteinene som er avhengige kalsium-calmodulin og blant dem dusinvis av enzymer, så som proteiner, proteinfosfase, nukleotid-syklasser og fosfodiesteraser; Alle involverte i utallige fysiologiske funksjoner som inkluderer:
- Metabolisme
- Partikkeltransport
- Visceral mobilitet
- Stoffsekresjon
- Eggløsning befruktning
- Genetisk uttrykk
- Celleproliferasjon
- Den strukturelle integriteten til celler
- Intercellulær kommunikasjon, etc.
Blant calmodulinavhengige proteiner er sitert: den lette kjeden Cinase of Myosin (MLCK), Cinase Phosphorylase og Ca ++/Calmodulin Cinasas I, II og III.
Dermed er informasjonen som er kodet av kalsiumsignalene (økning eller reduksjon i dens intracellulære konsentrasjon) "dechiffherd" av dette og andre kalsiumunionsproteiner, som gjør signalene til en biokjemiske endringer; Med andre ord, calmodulin er et mellomliggende protein i kalsiumavhengige signalprosesser.
Kan tjene deg: GLUT 2: Kjennetegn, struktur, funksjonerVirkningsmekanismen
Calmodulin er et veldig allsidig protein, ettersom dets "mål" -proteiner er betydelig forskjellige i form, sekvens, størrelse og funksjon. Siden det er et protein som fungerer som en "sensor" av kalsiumioner, avhenger virkningsmekanismen av endringene som er indusert i dens struktur og/eller konformasjon når den er sammen med fire av disse ionene.
Handlingsmekanismene kan eksemplifiseres ved kort å gjennomgå deres deltakelse i et par fysiologiske prosesser som sammentrekning av visceral glatt muskel og tilpasning til luktene som nescellene i luktslimhinnen har lert.
Calmodulin og glatt muskelkontraksjon
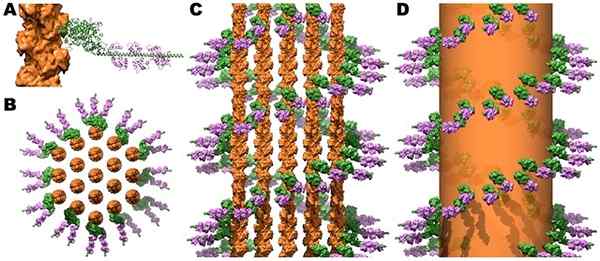 Struktur av myosin korsbruker og calmodulin i mikrovelloserende aktinstråler. Kilde: Jeffrey W. Brown, c. James McKnight [CC av (https: // creativecommons.Org/lisenser/av/3.0)]
Struktur av myosin korsbruker og calmodulin i mikrovelloserende aktinstråler. Kilde: Jeffrey W. Brown, c. James McKnight [CC av (https: // creativecommons.Org/lisenser/av/3.0)] Sammentrekningen av skjelett- og hjertemusklene utløses når økningen i cytosolisk Ca ++ når nivåer over 10-6 mol/L og dette ionet binder seg til troponin C, som gjennomgår endringer i de osteriske som påvirker tropomiosin. På sin side beveger tropomiosin seg og avslører i aktinforeningene med myosin, og stammer at den kontraktile prosessen utløser.
I den glatte muskelen er det ingen troponin C, og økningen av Ca ++ over det angitte nivået fremmer sin forening med calmodulin. Ca-Calmodulin-komplekset som er aktivt til den lette kjeden av myosin (MLCK), som igjen fosforylerer til denne lette kjeden, aktiverer myosin og utløser den kontraktile prosessen.
Økningen i Ca ++ er gitt ved inngangen fra utsiden eller avkjørsel fra det sarkoplasmatiske retikulum ved handling av inositol -tryfosfat (IP3) frigitt av fosfolipase C i fossen aktivert av GQ -proteinkoblede reseptorer. Avslapning oppstår når Ca ++, ved transporthandling, fjernes fra cytosol og går tilbake til kildestedene.
En viktig forskjell mellom begge typer sammentrekning er at CA ++ i de stripete musklene (hjerte og skjelett) induserer alestheriske forandringer når de blir sammen med proteinet, troponinet, mens de glatte endringene produsert av Ca-calmodulin er kovalent og antyder antyde myosinfosforylering.
Derfor, når Ca ++ -handlingen er ferdig, er deltakelse av et annet enzym som fjerner fosfat tilsatt av CINASA påkrevd. Dette nye enzymet er fosfatase av den lette kjeden til myosin (MLCP) hvis aktivitet ikke er avhengig av calmodulin, men er regulert av andre veier.
Faktisk opphører den kontraktile glatte muskelprosessen ikke, men sammentrekningsgraden opprettholdes på et mellomnivå, noe reguleringskontroller.
Kan tjene deg: pachyneTilpasning i luktesensorer
Den luktige sensasjonen utløser når luktreseptorer som ligger i cilia av celler som ligger på overflaten av luktslimhinnen, aktiveres.
Disse reseptorene er koblet til et heterotroft G -protein kjent som "golf" (G olfactory protein), som har tre underenheter: "αolf", "ß" og "γ".
Når luktreseptorene aktiveres som respons på en lukt, blir underenhetene til dette proteinet dissosiert og underenheten "αolf" aktiverer adenylciclase -enzymet, og produserer adenosin -syklisk monofosfat (AMPC).
AMPC aktiverer CNG -type kanaler (aktivert av sykliske nukleotider) for kalsium og natrium. Disse ionene kommer inn.
Kalsium som kommer inn, som har en tendens til å depolarisere cellen, har en antagonistisk effekt av negativ tilbakemelding, noe senere, ved å slå sammen calmodulinet og mellom dem som lukker kanalen og eliminerer depolariserende stimulus, til tross for at den luktige stimulansen vedvarer. Dette kalles sensortilpasning.
Calmodulin i planter
Planter reagerer også på intracellulære konsentrasjonsforskjeller fra kalsiumioner ved hjelp av calmodulinprotein. I disse organismer deler calmodulins mange strukturelle og funksjonelle egenskaper med sine kolleger hos dyr og gjær, selv om de er forskjellige i noen funksjonelle aspekter.
For eksempel blir calmodulin i planter sammen med korte peptidsekvenser i sine hvite proteiner, noe som induserer strukturelle endringer som endrer deres aktiviteter som svar på interne kalsiumvariasjoner.
I hvilken grad calmodulin kontroller i planter analoge prosesser som de forekommer til dyr, er noe som fremdeles er gjenstand for diskusjon i dag.
Referanser
- Brenner B: Muskulatur, i: Fysiologi, 6. utg; R Klinke et al (Eds). Stuttgart, Georg Thieme Verlag, 2010.
- Ganong WF: Molekylær basis for medisinsk fysiologi, i: i: Gjennomgang av medisinsk fysiologi, 25. utg. New York, McGraw-Hill Education, 2016.
- Guyton AC, Hall JI: Introduksjon til endokrinologi, i: Lærebok for medisinsk fysiologi, 13. utg, AC Guyton, JE Hall (eds). Philadelphia, Elsevier Inc., 2016.
- Kaup UB, Müller F: Olfactorisches System, i: Physiologie, 4. utg; P Deetjen et al (eds). München, Elsevier GmbH, Urban & Fischer, 2005.
- Korbmacher C, Greger R, Brenner B, Silbernagl S: Die Zelle Als Grundbautein, IN: Fysiologi, 6. utg; R Klinke et al (Eds). Stuttgart, Georg Thieme Verlag, 2010.
- Zielinski, r. OG. (1998). Calmodulin og calmodulin-bindende proteiner i planter. Årlig gjennomgang av plantebiologi, 49 (1), 697-725.