

Ikke -Mendelian kulturmønstre og eksempler
- 1405
- 379
- Theodor Anders Hopland
Med "Ikke -Mendelian arv”Vi viser til ethvert arvemønster der arvelige karakterer ikke skiller ut i samsvar med bestemmelsene i Mendels lover.
I 1865 laget Gregor Mendel, som ble ansett som "Genetics far", en serie eksperimentelle kryss med erteplanter, hvis resultater førte til at han foreslo postulater (Mendels lover) som forsøkte å gi en logisk forklaring til arv til arven etter karakterer mellom foreldre og barn.
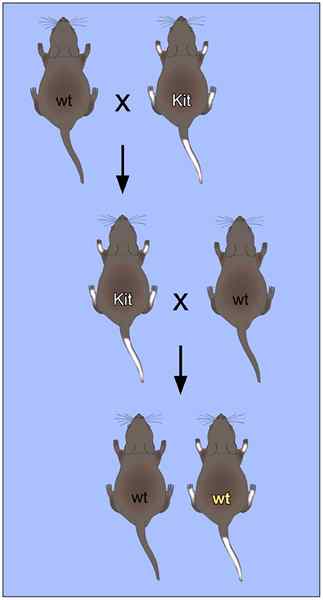
Ikke-Mendelian arv ved ville mus krysser og mutanter for benfenotypen og hvit halen (Kilde: Reinhard Liebers, Mineoo Rassoulzadegan, Frank Lyko [CC BY-S (https: // CreativeCommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Denne østerrikske munkemonken. I tillegg bestemte det de matematiske mønstrene som beskrev arven fra en generasjon til en annen, og disse funnene ble "beordret" i form av 3 grunnleggende lover:
- Dominansloven
- Loven om segregering av karakterer og
- The Law of Independent Distribution.
Suksessene og fradragene til Mendel ble skjult i mange år, inntil deres gjenoppdagelse på begynnelsen av 1900 -tallet.
 Gregor Mendel, betraktet som genetikkens far. Kilde: av Bateson, William (Mendels arvelighet: A Defense) [Public Domain], via Wikimedia Commons
Gregor Mendel, betraktet som genetikkens far. Kilde: av Bateson, William (Mendels arvelighet: A Defense) [Public Domain], via Wikimedia Commons På den tiden beholdt imidlertid det vitenskapelige samfunnet en noe skeptisk posisjon angående disse lovene, siden de ikke så ut til å forklare de arvelige mønstrene i noe dyre- eller plantearter, spesielt i de karakterene bestemt av mer enn ett lokus.
På grunn av dette klassifiserte de første genetikerne de arvelige mønstrene som ble observert som "Mendelians" (de som kunne forklares gjennom segregeringen av enkle, dominerende eller recessive alleler, som tilhører samme sted) og "ikke Mendelians" (de som ikke gjør de kan forklares så lett).
[TOC]
Ikke -Mendelian arvemønstre
Mendelian -arven refererer til et arvelig mønster som er i samsvar med lovene om segregering og uavhengig distribusjon, i henhold til hvilket et gen arvet fra alle foreldres segregater i gameter med en tilsvarende frekvens eller snarere, med samme sannsynlighet.
Hovedmønstrene for Mendelian arv som er beskrevet for noen sykdommer er: autosomal recessiv, autosomal dominerende og knyttet til X -kromosomet, som gir dominansmønstrene og resessiviteten beskrevet av Mendel.
Imidlertid ble disse postulert med hensyn til synlige trekk og ikke til gener (det bør tas i betraktning at noen alleler kan kode egenskaper som skiller ut som dominerende, mens andre kan kode de samme funksjonene, men de skiller ut som recessive gener).
Fra det ovennevnte følger det at den ikke -mendeliske arven bare består i ethvert arvelig mønster som ikke samsvarer med normen der et gen som er arvet fra ethvert foreldrenes segregue i cellene i kimlinjen med de samme sannsynlighetene og blant disse er inkludert:
Kan tjene deg: DNA -oversettelse: Eukaryot og prokaryotes prosess- Mitokondriell arv
- ""Avtrykk""
- Uniparental dysomi
- Ufullstendig dominans
- Kodominans
- Flere alleler
- Pleiotropy
- Dødelige alleler
- Polygenfunksjoner
- Arv knyttet til sex
Forekomsten av disse variasjonene i arvelige mønstre kan tilskrives de forskjellige interaksjonene som gener har med andre cellekomponenter, i tillegg til det faktum at hver og en er underlagt regulering og variasjon i noen av transkripsjonsstadiene, skjæring og skjøting, oversettelse , proteinfolding, oligomerisering, translokasjon og avdeling i cellen og for eksport.
Det er med andre ord mange epigenetiske påvirkninger som kan endre de arvelige mønstrene for enhver egenskap, noe som resulterer i et "avvik" fra Mendels lover.
Mitokondriell arv
Mitokondriell DNA overfører også informasjon fra en generasjon til en annen, akkurat som den som er inneholdt i kjernen til alle eukaryote celler gjør. Genomet som er kodet i dette DNA inkluderer de nødvendige genene for syntese av 13 polypeptider som er en del av underenhetene i den mitokondrielle respirasjonskjeden, essensielt for organismer med aerob metabolisme.
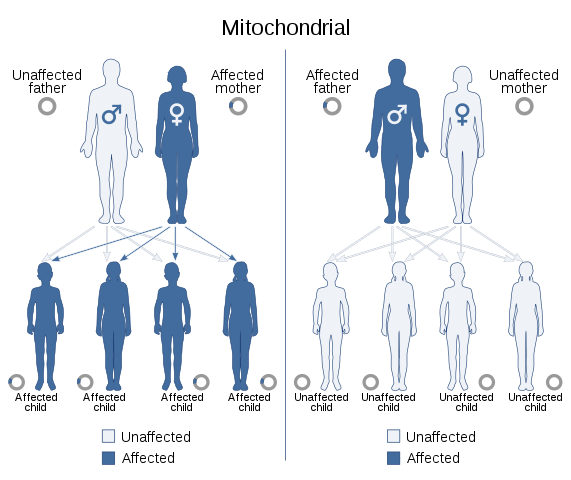 Mitokondrielle arvemønstre der begge foreldrene kan påvirkes (kilde: fil: selv -auto dominerende - i.SVG: Domaina, Angelito7 og SUM1Derivativt arbeid: SUM1 [CC BY-S (https: // CreativeCommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Mitokondrielle arvemønstre der begge foreldrene kan påvirkes (kilde: fil: selv -auto dominerende - i.SVG: Domaina, Angelito7 og SUM1Derivativt arbeid: SUM1 [CC BY-S (https: // CreativeCommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) De funksjonene som er resultatet av mitokondriell genom viser et spesifikt segregeringsmønster som har blitt kalt "mitokondriell arv", som vanligvis skjer gjennom mors linje, siden eggløsningen gir det totale komplementet av mitokondriell DNA og ingen mitokondrier er bidratt av sædspermen.
""Avtrykk"" eller genavtrykk
Det genomiske avtrykket består av en serie epigenetiske "merker" som kjennetegner visse komplette gengener eller regioner som følger av genomisk transport til hannen eller hunnen ved prosessen med gametenese.
Det er gengruppering, som består mellom 3 og 12 gener fordelt mellom 20 og 3700 kilo DNA -baser. Hver gruppe har en region kjent som avtrykkskontrollregionen, som viser spesifikke epigenetiske modifikasjoner fra hver foreldre blant dem er inkludert:
- DNA -metylering på spesifikke alleler i CpG -par cytokinavfall
- Post -translasjonell modifisering av histoner relatert til kromatin (metylering, acetylering, fosforylering, etc., av aminosyrehalene til disse proteinene).
Begge typer "merker" modulerer permanent uttrykket av genene de er, og endrer overføringsmønstrene til neste generasjon.
Kan tjene deg: DNA -emballasjeArvmønstre der uttrykket av en sykdom avhenger av de spesifikke allelene som er arvet fra en av de to foreldrene, er kjent som effekten av foreldrenes opprinnelse.
Uniparental dysomi
Dette fenomenet er et unntak fra Mendels første lov, som slår fast at bare en av de to allelene som er til stede i hver foreldre blir overført til avkom neste generasjon.
Det er et unntak fra regelen siden Uniparental Disomy er arven etter begge kopier av et homologt kromosom av en av foreldrene. Denne typen arvelige mønster viser ikke alltid fenotypiske defekter, med tanke på at den opprettholder de numeriske og strukturelle egenskapene til diploide kromosomer.
Ufullstendig dominans
Dette arvelige mønsteret består fenotypisk sett, i en blanding av funksjonene som er kodet av alleler som er kombinert. I tilfeller av ufullstendig dominans viser de individer som er heterozygot en blanding av funksjonene fra de to allelene som kontrollerer dem, noe som innebærer at forholdet mellom fenotypene er modifisert.
Kodominans
Beskriv de arvelige mønstrene der de to allelene som overføres fra foreldre til barna sine, uttrykker seg samtidig hos de som har heterozygot fenotype, så begge blir ansett som "dominerende".
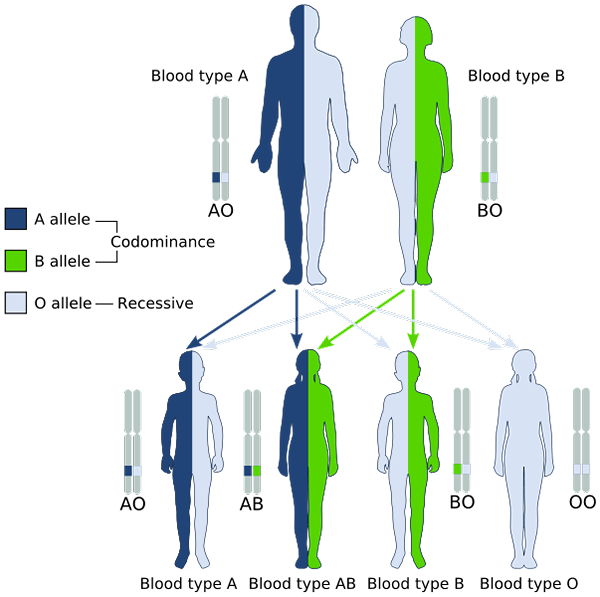 Eksempel på kodominans i ABO-systemet med blodgrupper (kilde: gyssinemraboTalk✉ Dette W3C-usikifiserte bildet ble opprettet med Inkscape. [Public Domain] via Wikimedia Commons)
Eksempel på kodominans i ABO-systemet med blodgrupper (kilde: gyssinemraboTalk✉ Dette W3C-usikifiserte bildet ble opprettet med Inkscape. [Public Domain] via Wikimedia Commons) Med andre ord, den recessive allelen er ikke "maskert" av uttrykket av den dominerende allelen i det alleliske paret, men begge uttrykkes og observeres, i fenotypen, en blanding av de to funksjonene.
Flere alleler
Allelene til et gen (kilde: Thomas Splettstoesser [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Kanskje er en av de viktigste svakhetene ved den Mendelian arven representert av funksjonene som er kodet av mer enn ett allel, som er ganske vanlig hos mennesker og i mange andre levende vesener.
Dette arvelige fenomenet øker mangfoldet av funksjonene som er kodet av et gen, og i tillegg kan disse genene også oppleve mønstre av ufullstendig dominans og kodominans i tillegg til enkel eller fullstendig dominans.
Pleiotropy
En annen av "steinene i skoen" eller "løse ben" fra Mendels arvelige teorier har å gjøre med de genene som kontrollerer utseendet til mer enn en fenotype eller synlig karakteristikk, som tilfellet er pleiotropiske gener.
Dødelige alleler
I sine verk vurderte Mendel heller ikke arven etter visse alleler som kan forhindre overlevelse av avkom når han er i den homozygot eller heterozygotform; Dette er dødelige alleler.
Det kan tjene deg: hva er en apomorfi? (Med eksempler)Dødelige alleler er vanligvis relatert til mutasjoner eller defekter i gener som er strengt nødvendige for å overleve, som når de overføres til neste generasjon (disse mutasjonene), avhengig av homozygose eller heterozygose hos individer, er dødelige.
Polygenegenskaper eller arv
Det er egenskaper som styres av mer enn ett gen (med deres alleler), og som i tillegg er sterkt kontrollert av miljøet. Hos mennesker er dette ekstremt vanlig, og det er tilfelle egenskaper som høyde, øyne, hår og hud, samt risikoen for å lide av noen sykdommer.
Arv knyttet til sex
Hos mennesker og mange dyr er det også egenskaper som finnes i et av de to kjønnskromosomene, og som overføres gjennom seksuell reproduksjon. Mange av disse funksjonene anses som "sexkoblet" når de er bevist i et av kjønnene, selv om de to er fysisk i stand til å arve slike funksjoner.
De fleste av de kjønnsrelaterte funksjonene er assosiert med noen recessive sykdommer og lidelser.
Eksempler på ikke -Mendelian arv
Det er en genetisk lidelse hos mennesker kjent som elfenbenssyndrom, som er forårsaket av en mutasjon i et enkelt gen som samtidig påvirker vekst og utvikling (høyde, syn og hjertefunksjon, blant andre).
Dette er en sak som et utmerket eksempel på det ikke -Mendeliske arvemønsteret kalt Pleiotropia, der et enkelt gen kontrollerer forskjellige egenskaper.
Mitokondriell arveeksempel
De genetiske lidelsene som følger av mitokondrisk DNA har utallige kliniske fenotypiske variasjoner siden det som er kjent som heteroplasmi, der forskjellige vev har en annen prosentandel av mutant mitokondrielt genom og derfor presenterer forskjellige fenotyper.
Blant disse lidelsene er "utmattelse" -syndromene til mitokondrier, som er en gruppe autosomale recessive lidelser som er preget av en viktig reduksjon i mitokondriell DNA -innhold, som ender med mangelfulle energiproduksjonssystemer i de mest berørte organene og vevene.
Disse syndromene kan skyldes mutasjoner i det nukleære genomet som påvirker de nukleære genene som er involvert i mitokondrielle nukleotider -syntese eller i replikasjonen av mitokondriell DNA. Effektene kan bevises som myopatier, encefalopatier, hepato-cerebral eller neuro-gastrointestinal defekter.
Referanser
- Gardner, J. OG., Simmons, J. OG., & Snustad, d. P. (1991). Hoved av genetisk. 8 '"utgave. Jhon Wiley og sønner.
- Griffiths, a. J., Wessler, s. R., Lewontin, r. C., Gelbart, w. M., Suzuki, d. T., & Miller, J. H. (2005). En introduksjon til genetisk analyse. Macmillan.
- Harel, t., Pehlivan, d., Caskey, c. T., & Luppski, J. R. (2015). Mendelian, ikke-Mendelian, multigenarv og epigenetikk. I Rosenbergs molekylære og genetiske grunnlag for nevrologisk og psykiatrisk sykdom (PP. 3-27). Akademisk presse.
- Sølv, l. (2001). Ikke-Mendelian arv.
- Van Heyningen, V., & Yeyati, s. L. (2004). Mekanismer for ikke-kjelig arv ved genetisk sykdom. Human Molecular Genetics, 13 (Suppl_2), R225-R233.
- « Hvordan reproduserer planter? Aseksuell og seksuell reproduksjon
- Calmodulin Structure, Functions and Mechanism of Action »