

Kjennetegn, histologi, funksjoner, dyrkingskondrocytter

- 4623
- 1301
- Mathias Aas
De Kondrocytter De er de viktigste bruskcellene. De er ansvarlige for sekresjonen av den ekstracellulære matrisen til brusk, dannet av glukosaminoglykaner og proteoglykaner, kollagenfibre og elastiske fibre.
Brusk er en spesiell type konjunktivt vev av hvitaktig, resistent og elastisk farge som danner skjelettet eller tilsettes visse bein av noen virveldyr.
 Kutt av et bruskvev, nummer 2 indikerer plasseringen av en kondrocytt (kilde: Guido furapani [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Kutt av et bruskvev, nummer 2 indikerer plasseringen av en kondrocytt (kilde: Guido furapani [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Brusken hjelper også til å forme flere organer som nese, ører, larynx og andre. I henhold til typen fibre som er inkludert i den hemmelige ekstracellulære matrisen, er bruskene klassifisert i tre typer: (1) hyalinbrusk, (2) elastisk brusk og (3) fibrocartílago.
De tre typene brusk har to vanlige bestanddeler: celler, som er kondroblaster og kondrocytter; og matrisen, dannet av fibre og et grunnleggende stoff som ligner på en gel som etterlater små områder kalt “laguner” der cellene er lokalisert.
Den brusk matrisen mottar ikke blodkar, lymfekar eller nerver og gir næring ved diffusjon fra det omkringliggende bindevevet eller, i tilfelle av synoviale ledd, fra synovialvæske.
[TOC]
Kjennetegn
Chondrocyttene er til stede i de tre brusktypene. De er celler avledet fra mesenkymale celler, som i områdene der brusk dannes, mister utvidelsene, er avrundet og samles og danner tette masser kalt sentre for "kondrifisering".
I disse kondrifiseringssentrene er stamceller forskjellige i kondroblaster, som begynner å syntetisere den bruskmatrisen som gradvis omgir dem.
Analog med hva som skjer med osteocytter (beinceller), kondroblastene som er inkludert i de så -kallede "lagunene" av matrisen, er forskjellige i kondrocytter.
Kondrocytter i hullene deres kan deles, og danner klynger på omtrent fire eller flere celler. Disse klyngene er kjent som isogene grupper og representerer de opprinnelige kondrocyttavdelingene.
Bruskvekst og kondroblastdifferensiering
I den grad hver celle i hver klynge eller isogen gruppe danner matrise, beveger de seg bort fra hverandre og danner sine egne separate hull. Som en konsekvens vokser brusk fra innsiden, og kaller denne formen for bruskvekst som mellomliggende vekst.
Kan tjene deg: trofoblastI de perifere regionene i den utviklende brusk, er mesenkymale celler forskjellige i fibroblaster. Disse syntetiserer et uregelmessig kollagenøst bindevev kalt Pericondrio.
Pericondrium har to lag: en ekstern vaskularisert fibrøs og sammensatt av type I kollagen og fibroblaster; og et annet indre cellelag dannet av kondrogene celler som er delt og differensierer i kondroblaster, som danner matrisen som er perifert tilsatt.
Gjennom denne differensieringen av perikondriumceller vokser også brusk ved perifer apposisjon. Denne vekstprosessen kalles apostal vekst.
Interstitiell vekst er typisk for den innledende fasen av bruskutvikling, men den forekommer også i leddbrusk som ikke har perikondrium og i epifysiske plater eller vekstplater i de lange beinene.
I resten av kroppen, derimot, vokser brusk ved apposisjon.
Histologi
I bruskene kan du finne tre typer konrogene celler: kondroblastene og kondrocyttene.
Kondrogene celler er tynne og langstrakte i spindel og stammer fra differensiering av mesenkymale celler.
Kjernen er ovoid, de har lite cytoplasma og et dårlig utviklet Golgi -kompleks, mitokondrier og grov endoplasmatisk retikulum. De kan differensiere i kondroblaster eller osteoprogenitorceller.
Kondrogene celler i det indre laget av perikondrium, så vel som de mesenkymale cellene i kondrifikasjonssentrene, er de to kildene til kondroblaster.
Disse cellene har en stor utvikling av grov endoplasmatisk retikulum, mange ribosomer og mitokondrier, et godt utviklet Golgi -kompleks og tallrike sekresjonsvesikler.
Kondrocytter i bruskvev
Kondrocytter er kondroblaster omgitt av ekstracellulær matrise. De kan ha en ovoid form når de er nær periferien, og en mer avrundet form med omtrent 20 til 30μm i diameter når de er i dypere regioner av brusk.
Unge kondrocytter har en stor kjerne med en fremtredende nukleolus og rikelig cytoplasmatiske organeller som Golgi -kompleks, grov endoplasmatisk retikulum, ribosomer og mitokondrier. De har også rikelig glykogencytoplasmatiske avsetninger.
Gamle kondrocytter har få organeller, men rikelig frie ribosomer. Disse cellene er relativt inaktive, men kan aktiveres ved å øke proteinsyntesen.
Kondrocytter og brusktyper
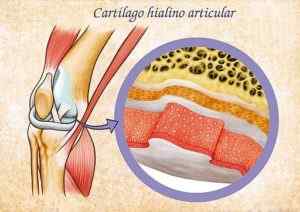
Arrangementet av kondrocyttene varierer i henhold til typen brusk der de er. I Hyalino -brusk, som har en perle og gjennomskinnelig hvitt utseende, danner kondrocyttene mange isogene grupper og anordnet i store hull med svært få fibre i matrisen.
Kan tjene deg: osteocytter: trening, egenskaper og funksjonerJoint Hyalino brusk (kilde: Eugenio Fernández Pruna [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) Hialino brusk er det mest tallrike i menneskets skjelett og inneholder kollagenfibre type II.
I den elastiske brusk, som har rikelig forgrenede elastiske fibre og sammenvevd med type II kollagenfibre fordelt over matrisen, er kondrocyttene rikelig og er jevnt fordelt mellom fibrene.
Denne typen brusk er typisk for den atriale paviljongen, Eustachian -rørene, noe laryngeal brusk og epiglottis.
I Fibrocartíla er det få condoches rettet mellom dens tykke og tett distribuerte type I kollagenfibre i matrisen.
Denne typen brusk er lokalisert på de intervertebrale skivene, i symfysen av pubis, i seneinnsettingsområdene og i kneleddet.
Funksjoner
Den grunnleggende funksjonen til kondrocytter er å syntetisere den ekstracellulære matrisen til de forskjellige typer brusk. Som kondrocytter, sammen med matrisen, er de de konstituerende elementene i brusk og deler deres funksjoner med den (som helhet).
Blant hovedfunksjonene til brusk er dem av demping eller absorberende støt eller slag og kompresjoner (takket være deres motstand og fleksibilitet).
I tillegg gir de en jevn artikulær overflate som tillater artikulære bevegelser med minimal friksjon og til slutt former forskjellige organer som atrial paviljong, nese, larynx, epiglottis, bronchi osv.
Avlinger
Hialino brusk, som er den mest tallrike av menneskekroppen, kan bli utsatt for flere sykdomsskader, men fremfor alt for sportspraksis.
Å være at brusk er et høyt spesialisert stoff, og med relativt lite selvreparasjonskapasitet kan skadene deres forårsake irreversibel skade.
Mange kirurgiske teknikker er utviklet for å reparere lesjonene i leddbrusken. Mens disse teknikkene, noen mer inngripende enn andre, kan forbedre lesjoner, dannes den reparerte brusk som en fibrocartílago og ikke som hyalinbrusk. Dette betyr at den ikke har de samme funksjonelle egenskapene som den opprinnelige brusk.
Det kan tjene deg: Flagelos: Eukaryota, Procariota (struktur og funksjoner)For å oppnå tilstrekkelig reparasjon av skadede artikulære overflater, er autologe avlingsteknikker utviklet (fra deres egen brusk) for å oppnå in vitro vekst av brusk og dens påfølgende transplantasjon.
Disse avlingene har utviklet isolering, en prøve av sunn brusk av pasienten, kondrocyttene som deretter dyrkes og transplanteres.
Disse metodene har vist seg å være effektive for vekst og utvikling av hyalin -leddbrusk, og etter en omtrentlig periode på to år, oppnå den definitive utvinningen av artikulær overflate.
Andre teknikker involverer bruskkultur In vitro på en matrise eller gel av fibrin og algininsyre eller andre naturlige eller syntetiske stoffer som er under studie som for øyeblikket er undersøkt.
Målet med disse avlingene er imidlertid å gi materiale for transplantasjon av skadde leddflater og deres definitive utvinning.
Referanser
- Doubek, r. W. (1950). Histologi med høyt avkastning (2. utg.). Philadelphia, Pennsylvania: Lippinott Williams & Wilkins.
- Gartner, l., & Hiatt, j. (2002). Histology Atlas Text (2. utg.). Mexico d.F.: McGraw-Hill Inter-American Editors.
- Giannini, s., R, b., Grigolo, f., & Vannini, f. (2001). Autolog kondrocyttransplantasjon i osteokondrale lesjoner av ankellleddet. Fot- og ankelinternasjon, 22(6), 513-517.
- Johnson, k. (1991). Histologi og cellebiologi (2. utg.). Baltimore, Maryland: The National Medical Series for Independent Study.
- Kino-oka, m., Maeda, og., Yamamoto, t., Sugawara, k., & Taya, m. (2005). En kinetisk modellering av kondrocyttkultur for fremstilling av vevs-konstruert brusk. Journal of Bioscience and BioEngineering, 99(3), 197-207.
- Park, og., Lutolf, m. P., Hubbell, J. TIL., Hunziker, e. B., & Wong, m. (2004). Bovint primær kondrocyttkultur i syntitisk matriks metalloproteina -sensitiv poly (etylenglykol) -baserte hydrogeler som et stillas for bruskreparasjon. Vevteknikk, 10(3-4), 515-522.
- Perka, c., Spitzer, r. S., Lindenhayn, k., Sitterende, m., & Schultz, eller. (2000). Matrix-blandet kultur: Ny metodikk for kondrocyttkultur og utarbeidelse av brusktransplantasjoner. Journal of Biomedical Materials Research, 49, 305-311.
- Qu, c., Puttonen, k. TIL., Lindeberg, h., Ruponen, m., Hovatta, eller., Koistinaho, J., & Lammi, m. J. (2013). Kondrogen differensiering av humane pluripotente stamceller i kondrocytt samkultur. International Journal of Biochemistry and Cell Biology, Fire fem, 1802-1812.
- Ross, m., & Pawlina, w. (2006). Histologi. En tekst og atlas med korrelert celle- og molekylærbiologi (5. utg.). Lippinott Williams & Wilkins.
- « Antiestreptolisin eller kvantifisering, fundament, teknikk, patologier
- Manguesta -egenskaper, habitat, reproduksjon og mat »