

Sfingolipider hva er, egenskaper, funksjoner, syntese
- 4429
- 952
- Prof. Oskar Aas
Hva er sphylingolipider?
De sfingolipider De representerer en av de tre store lipidfamiliene som er til stede i biologiske membraner. Som glyceophospholipider og steroler, er de amfipatiske molekyler med en hydrofil polarregion og en hydrofob apolar region.
De ble først beskrevet i 1884 av Johann L. W. Thudichum, som beskrev tre sfingolipider (sfingomyeline, hjerne og hjerne -brain) som tilhører de tre forskjellige klassene som er kjent: fosfosfosfingolipider, glykosfingolipider nøytrale og sure.
I motsetning til glyceofosfolipider, er ikke sphyingolipider bygget på et 3-fosfatglyserolmolekyl som hovedskjelettet, men er forbindelser avledet fra sfiny, en aminoalkohol med en lang hydrokarbonkjede festet av en Amida-binding.
OGStruktur av sfingolipidene
Alle sfingolipider stammer fra en L-serine, som er kondensert med en langkjede fettsyre for å danne sfingoidbasen, også kjent som Long Chain Base (LCB).
De vanligste basene er sfinganin og sfinxin, som skiller seg fra hverandre i nærvær av en dobbeltbinding trans Mellom karbonatomer 4 og 5 av fettsyren i sfinxinet.
Karbon 1, 2 og 3 i sfinxinet er strukturelt analogt med glyserol glyserofosfolipider glyserol. Når Amida bindinger binder en fettsyre til karbon 2 av sfinen.
Langkjedede fettsyrer som utgjør de hydrofobe regionene i disse lipidene kan være veldig mangfoldige. Lengdene varierer fra 14 til 22 karbonatomer som kan ha forskjellige metningsgrader, vanligvis mellom karbonatomer 4 og 5.
I posisjoner 4 eller 6 kan de ha hydroksiljegrupper og dobbeltbindinger i andre posisjoner eller til og med konsekvenser som metylgrupper.
Kjennetegn
Fettsyrekjedene knyttet av AMIDA -bindinger til ceramidene er ofte mettede, og har en tendens til å være av større lengde enn de som finnes i glyserofosfolipider, noe som ser ut til å være avgjørende for den biologiske aktiviteten til disse.
Et særegent kjennetegn ved sfingolipidskjelettet er at de kan ha en positiv nettbelastning til nøytral pH, rart blant lipidmolekyler.
Imidlertid PkA av aminogruppen er lav med hensyn til en enkel amin, mellom 7 og 8, så en del av molekylet er ikke lastet til fysiologisk pH, noe som kan forklare den "frie" bevegelsen til disse blant bilapas.
Den tradisjonelle klassifiseringen av sfingolipider oppstår fra de flere modifikasjonene som ceramidmolekylet kan lide, spesielt med tanke på erstatningene til polarehodegruppene.
Kan tjene deg: Hepadnavirus: Kjennetegn, morfologi, behandlingFunksjoner
Sphingolipider er essensielle hos dyr, planter og sopp, så vel som i noen prokaryote organismer og virus.
-Strukturelle funksjoner
Sphingolipider modulerer de fysiske egenskapene til membraner, inkludert deres fluiditet, tykkelse og krumning. Modulær disse egenskapene gir dem også direkte innflytelse på den romlige organiseringen av membranproteiner.
I lipid "flåter"
I biologiske membraner kan dynamiske mikrodomener oppdages med lavere flyt dannet av kolesterol og sfingolipidmolekyler kalt lipidbalsas.
Disse strukturene forekommer naturlig og opprettholder et nært forhold til omfattende proteiner, celleoverflatreseptorer og signaleringsproteiner, transportører og annet protein med glykosylfosfatidylinitol (GPI) anker (GPI).
-Signaleringsfunksjoner
De har funksjoner som signalmolekyler som fungerer som andre budbringere eller som utskilte ligander for celleoverflatreseptorer.
Som sekundære budbringere kan de delta i reguleringen av kalsiumhomeostase, cellevekst, tumorogenese og undertrykkelse av apoptose. I tillegg avhenger aktiviteten til mange integrerte og perifere membranproteiner av dens tilknytning til sfingolipider.
Mange intercellulære og celleinteraksjoner med omgivelsene er avhengige av eksponeringen av de forskjellige polare gruppene av sfingolipidene mot det ytre ansiktet til plasmamembranen.
Foreningen av glykosfingolipider og lektinas er avgjørende for assosiasjonen av myelin med aksoner, vedheft av nøytrofiler til endotelet, etc.
Av -produkter av metabolismen hans
De viktigste signaliserende sfingolipidene er langkjedede baser eller sfinxer og ceramider, så vel som deres fosforylerte derivater, så som 1-fosfat sphygosin.
Metabolismeproduktene fra mange sfingolipider aktiverer eller hemmer flere hvite downstants (kinaseproteiner, fosfatose og andre), som kontrollerer cellulær atferd så kompleks som vekst, differensiering og apoptose.
-Som membranreseptorer
Noen patogener bruker gloysfingolipider som reseptorer for å formidle inngangen til vertsceller eller for å levere virulensfaktorer til dem.
Det er vist at sphyglyipider deltar i flere cellehendelser som sekresjon, endocytose, kjemiotaxis, nevrotransmisjon, angiogenese og betennelse.
De er også involvert i membrantrafikk, så de påvirker internaliseringen av reseptorer, orden, bevegelse og fusjon av sekretoriske vesikler som respons på forskjellige stimuli.
Sfingolipidgrupper
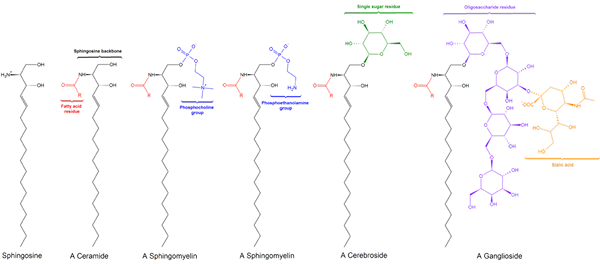
Det er tre sfingolipid underklasser, alle avledet fra ceramidet og som skiller seg fra hverandre av polargrupper, nemlig: sphylingomyelins, glykolipider og gangliasider.
Sphingomyielinas
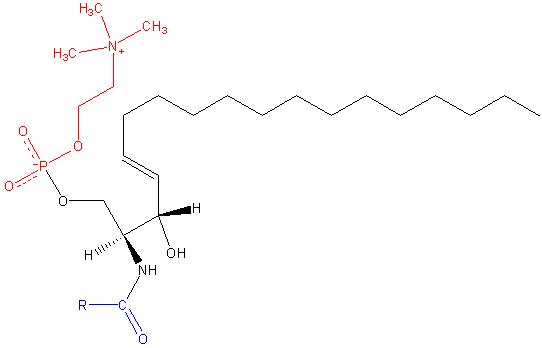 Sfingomilein. Svart: Sphinxine. Rødt: fosfokolin. Blå: fettsyre.
Sfingomilein. Svart: Sphinxine. Rødt: fosfokolin. Blå: fettsyre. Disse inneholder fosfokolin eller fosfoetanolamin som en polar hodegruppe, slik at de er klassifisert som fosfolipider sammen med glyceofosfolipider. De ser selvfølgelig fosfatidylkoliner i tre -dimensjonal struktur og generelle egenskaper siden de ikke har noen ladning i polare hoder.
Kan tjene deg: enolase: struktur, virkningsmekanisme, funksjonerDe er til stede i plasmamembranene i dyreceller og er spesielt rikelig i myelin, en pod som omgir og isolerer aksonene til noen nevroner.
Glykolipider eller nøytrale glykosfingolipider (uten belastning)
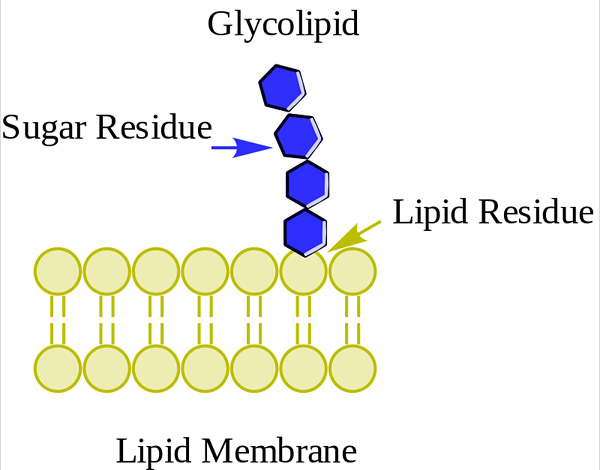 Glukolipid. WPCROSSON [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons
Glukolipid. WPCROSSON [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons De er først og fremst på det ytre ansiktet av plasmamembranen og har ett eller flere sukkerarter som en polarhodegruppe direkte forent til karbonhydroksyl 1 av ceramiddelen. De har ikke fosfatgrupper. Siden pH 7 ikke har noen belastning, kalles de nøytrale glykolipider.
Cerebrosider har et enkelt sukkermolekyl knyttet til ceramid. De som inneholder galaktose er i plasmamembraner av ikke -nerve vevsceller. Globosider er goustoesphingolipider med to eller flere sukkerarter, normalt d-glycose, d-galaktose eller n-acetyl-d-galaktosamin.
Gangliósidos eller glykosfingolipider
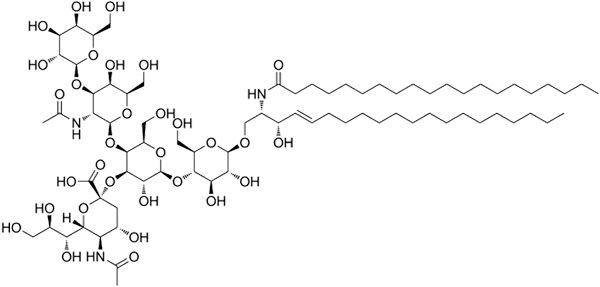 GM1 ganglosidstruktur
GM1 ganglosidstruktur Dette er de mest komplekse sfingolipider. De har oligosakkarider som en polar hodegruppe og en eller flere terminal N-acetylmuraminsyrerester, også kalt siralyre. Siamic acid bringer ganglia den negative belastningen til pH 7, som skiller dem fra nøytrale glykosfingolipider.
Nomenklaturen av denne typen sfingolipider avhenger av mengden sinalsyreavfall som er til stede i oligosakkariddelen av polarhodet.
Syntese
Det lange sjakten eller sphiny base molekylet syntetiseres i endoplasmatisk retikulum (ER) og tilsetningen av polargruppen i spissen for disse lipidene forekommer senere i Golgi -komplekset. Hos pattedyr kan noe syntese av sfingolipider også forekomme i mitokondrier.
Etter å ha fullført syntesen i Golgi -komplekset, blir sfingolipidene transportert til andre cellekommer gjennom mekanismer mediert av vesikler.
Biosyntesen av sfingolipidene består av tre grunnleggende hendelser: syntesen av de langkjedebasene, biosyntesen av ceramidene ved foreningen av en fettsyre gjennom en AMIDA -binding, og til slutt dannelsen av de komplekse sfingolipidene ved hjelp av unionen av polare grupper i karbon 1 av sfingoidebasen.
I tillegg til syntese av novo, Sphingolipider kan også dannes ved erstatning eller resirkulering av langkjedede baser og ceramider, som kan mate sfingolipidbassenget.
Ceramid -skjelettsyntese
Biosyntesen av ceramiden, skjelettet til sfingolipidene, begynner med den dekarboksylerative kondensasjonen av et palmitail-CoA-molekyl og en L-serin. Reaksjonen katalyseres av en serin palmitail transferase (SPT) heterodimerisk, pyridoksal fosfatavhengig, og produktet er 3-ZO dihydroesfingosin ZE.
Kan tjene deg: mikrobiota: komposisjon, funksjoner og hvor er detDette enzymet blir hemmet av ß-phaal-L-alaniner og L-cicloserinas. I gjær er det kodet av to gener, mens det i pattedyr er tre gener for dette enzymet. Det aktive stedet er i det cytoplasmatiske ansiktet til endoplasmatisk retikulum.
Rollen til dette første enzymet er bevart i alle de studerte organismer. Imidlertid er det noen forskjeller mellom taxa som har å gjøre med den subcellulære plasseringen av enzymet: bakterier er cytoplasmatisk, gjær, planter og dyr er i endoplasmatisk retikulum.
3-Zoesfinganin blir deretter redusert med NADPH-avhengig 3-ZO-ketosfinganin. Dihydroceramidsyntase (sfinganin N-acil transferase) acetyla deretter sfinanin for å produsere dihydroceramid. Ceramid blir deretter dannet av dihydroceramid deaturase/reduktase, som setter inn en dobbeltbinding trans I stilling 4-5.
Hos pattedyr er det mange ceramidisoformer syntaser, som hver sammenføyer en spesifikk fettsyrekjede til basene til lange kjeder. Derfor gir ceramidesynthesas og andre enzymer, Elongasas, den viktigste kilden til mangfold av fettsyrer i sfingolipider.
Metabolisme
Nedbrytningen av sfingolipidene utføres av glykohydroller og sfingomyelinase -enzymer, som er ansvarlige for å fjerne modifikasjonene av polargruppene. På den annen side regenererer ceramidasene de lange kjedebasene fra ceramidene.
Gangliene blir nedbrutt av et sett med lysosomale enzymer som katalyserer trinnet for trinnet til sukkerenhetene, og produserer en ceramid til slutt.
En annen måte å nedbryte på er internalisering av sfingolipider i endocytiske vesikler som blir sendt tilbake til plasmamembranen eller transportert til lysosomer der de blir nedbrutt av spesifikk syrehydrolase.
Ikke alle langkjedede baser blir resirkulert, endoplasmatisk beholdes. Denne nedbrytningsmekanismen består av fosforylering i stedet for acilatisering av LCB -ene, og gir opphav til signalmolekyler som kan være oppløselige underlag for LIASes -enzymet.
Regulering
Metabolismen til disse lipidene er regulert på flere nivåer, en av dem er den av enzymene som har ansvaret for syntese, deres post -translasjonelle modifikasjoner og de samme mekanismene av samme.
Noen reguleringsmekanismer er cellespesifikasjoner, enten for å kontrollere tiden for celleutvikling de produseres eller som svar på spesifikke signaler.
Referanser
- Bartke, n., & Hannun, og. (2009). Bioaktive sppingolipider: metabolisme og funksjon. Journal of Lipid Research, femti, 19.
- Breslow, d. K. (2013). Sfingolipid homeostase i endoplasmatisk retikulum og utover. Kald vårhavnperspektiver i biologi, 5(4), A013326.