

Transkripsjonsfaktor virkningsmekanisme, typer, funksjoner
- 2747
- 575
- Prof. Joakim Johansen
EN Transkripsjonsfaktor Det er et regulatorisk "tilbehør" protein som er nødvendig for genetisk transkripsjon. Transkripsjon er det første trinnet i genetisk uttrykk og innebærer overføring av informasjon i DNA til et RNA -molekyl, som deretter behandles for å gi opphav til genprodukter.
RNA -polymerase II er enzymet som har ansvaret for transkripsjonen av de fleste eukaryote gener og produserer, i tillegg til noen små RNA -er, messenger -RNA -ene som deretter vil bli oversatt til proteiner. Dette enzymet krever tilstedeværelse av en type transkripsjonsfaktorer kjent som generelle eller basale transkripsjonsfaktorer.
Dette er imidlertid ikke de eneste transkripsjonsfaktorene som eksisterer i naturen, siden det er "ikke-generelle" proteiner, både i eukaryoter og i prokaryoter og buer, som er involvert i reguleringen av den vevsspesifikke genetiske transkripsjonen (i flercellulære organismer ) eller i reguleringen av genaktivitet som respons på forskjellige stimuli.
Disse transkripsjonsfaktorene er effektorer av stor betydning, og kan finnes praktisk talt i alle levende organismer, siden de representerer hovedkilden til regulering av genetisk uttrykk.
Detaljerte studier av forskjellige transkripsjonsfaktorer i forskjellige typer levende organismer indikerer at de har en modulær struktur, der en spesifikk region er ansvarlig for interaksjon med DNA, mens andre produserer de stimulerende eller hemmende effektene.
Transkripsjonsfaktorene deltar da i modelleringen av de genetiske ekspresjonsmønstrene som ikke har noe å gjøre med endringene i DNA -sekvensen, men med epigenetiske endringer. Vitenskapen som er ansvarlig for studiet av disse endringene er kjent som epigenetikk.
[TOC]
Virkningsmekanismen
For å utøve sine funksjoner, må transkripsjonsfaktorer kunne gjenkjenne og bli med spesifikt en bestemt DNA -sekvens for å positivt eller negativt påvirke transkripsjonen av nevnte region av DNA.
De generelle transkripsjonsfaktorene, som i utgangspunktet er de samme for transkripsjonen av alle type II -gener i eukaryoter, er først samlet på promoterregionen til genet, og dirigerer dermed posisjoneringen av polymeraseenzymet og "åpningen" av dobbelt propell.
Prosessen er gitt av flere påfølgende trinn:
- Union av den generelle transkripsjonsfaktoren tfiid til en sekvens av gjentatt timina (t) og adenin (a) i genet kjent som “boksen Tata”; Dette forårsaker DNA -forvrengning som er nødvendig for forening av andre proteiner til promoterregionen.
- Posterior montering av andre generelle faktorer (tfiiB, tfiih, tfih, tfiie, tfiif, etc.) og av RNA -polymerase II, og danner det som kalles Transkripsjonsinitieringskompleks.
Kan tjene deg: kromatider- Liberasjon av initieringskomplekset, fosforylering av polymerase av TFIIH -faktoren og begynnelsen av transkripsjonen og syntesen av et RNA -molekyl fra sekvensen til genet som er transkribert.
Aktivering og undertrykkelse av transkripsjon
Som nevnt kan "ikke -generelle" transkripsjonsfaktorer regulere uttrykket av gener, enten det er positivt eller negativt.
Aktivering
Noen av disse proteinene inneholder, i tillegg til de strukturelle domenene til DNA -forening, andre grunner kjent som aktiveringsdomener, som er rike på sure syrerester, glutamin- eller prolinrester.
Disse aktiveringsdomenene samhandler med elementene i det generelle transkripsjonsfaktorkomplekset eller med beslektede koaktiverende molekyler som direkte samhandler med komplekset. Denne interaksjonen resulterer i stimulering av montering av transkripsjonskomplekset eller i økningen i aktiviteten.
Undertrykkelse
De fleste transkripsjonsfaktorer hemmer transkripsjon når du forstyrrer aktiviteten til transkripsjonsfaktorer som virker positivt, og blokkerer dens stimulerende effekt. De kan jobbe ved å blokkere foreningen av den positive faktoren til DNA eller virker på faktorene som inaktiverer strukturen til kromatinet.
Andre hemmende faktorer virker direkte å blokkere transkripsjonen, uten å blokkere virkningen av noen aktiverende transkripsjonsfaktor; og redusere basalnivået på transkripsjon, til et enda lavere nivå enn det oppnås i fravær av aktiverende faktorer.
Som å aktivere proteiner, fungerer undertrykkende faktorer direkte eller indirekte med basale eller generelle transkripsjonsfaktorer.
Folkens
Selv om de fleste transkripsjonsfaktorer er klassifisert i henhold til egenskapene eller identiteten til deres DNA -unionsdomener, er det noen, også klassifisert som transkripsjonsfaktorer, som ikke samhandler direkte med DNA og er kjent som transkripsjonsfaktorer "indirekte".
Direkte transkripsjonsfaktorer
De er de vanligste transkripsjonsfaktorene. De har DNA -unionsdomener og kan aktivere eller hemme genuttrykk ved hjelp av deres union til spesifikke DNA -regioner. De skiller seg fra hverandre, spesielt med hensyn til deres DNA -unionsdomener og deres oligomeriseringstilstand.
De mest studerte og anerkjente familiene av denne typen faktorer er:
Helix-Gutero-Hélice ("Helix-Turn-helix”, Hth)
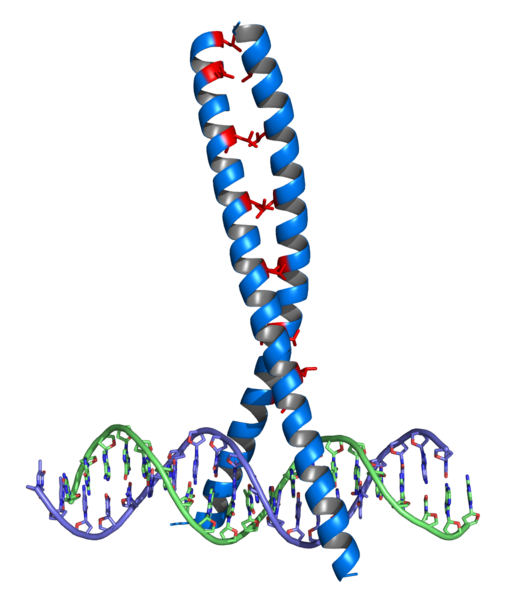
Dette var den første familien av faktorer med DNA Union -domener som ble oppdaget, og er til stede i mange eukaryoter og prokaryoter. Årsaken til anerkjennelse består av en α -propell, en sving og en annen propell α.
De har bevart domener av glycin i området for svingen, og også noe hydrofobt avfall som hjelper til med å stabilisere arrangementet av de to propellene i HTH -enheten.
Kan tjene deg: genetisk bassengHomeodominium
Det er til stede i et stort antall eukaryote regulatoriske proteiner. De første sekvensene ble gjenkjent i regulatoriske proteiner av utviklingen av Drosophila. Dette domenet inneholder en Hth-grunn til å slå sammen DNA og en ekstra α-propell, i tillegg til en utvidet N-terminal arm.
Sinkfingre
Ble oppdaget i transkripsjonsfaktoren tfiiia av Xenopus Og det har blitt vist at de deltar i mange aspekter av eukaryotisk genetisk regulering. De finnes i proteiner indusert av signaler om differensiering og vekst, i protooncogenes og i noen generelle transkripsjonsfaktorer.
De er preget av tilstedeværelsen av repetisjoner i en gruppe av motivene til sink på 30 avfall som inneholder flere avfall og histidinavfall.
Steroidreseptorer
Denne familien inkluderer betydelige regulatoriske proteiner som, i tillegg til å ha et domene for forening av hormoner, har et domene av DNA -union og vanligvis fungerer som transkripsjonsaktivatorer.
Unionsdomener inneholder 70 avfall, blant dem 8 er beholdt cysteinrester. Noen forfattere vurderer at disse faktorene kan danne et par sinkfingre, gitt tilstedeværelsen av to fire -Cystein -spill.
Leucina og helisk-bukle-stenging ("Helix-loop-helix ")
Disse transkripsjonsfaktorene er involvert i differensiering og utvikling og arbeid for dannelse av en heterodimer. Leucin Closure -domenet er observert i forskjellige eukaryoter og er preget av to underdomener: stenging av leuciner som måler dimerisering og et grunnleggende område for forening med DNA.
Årsaker β grunner
De finnes hovedsakelig i eukaryote faktorer og skiller.
Indirekte transkripsjonsfaktorer
Denne typen transkripsjonsfaktorer utøver sine regulatoriske effekter på genetisk uttrykk ikke gjennom dets direkte interaksjon med DNA, men gjennom protein-protein-interaksjoner med andre transkripsjonsfaktorer som samhandler med DNA. Det er grunnen til at de kalles "indirekte".
Den første som beskrev var trans-aktivering av viruset til "Herpes enkel " (HSV) kjent som VP16, som binder seg til OCT-1-faktoren når cellene er infisert med dette viruset, og stimulerer transkripsjonen av et spesifikt gen.
Faktorene av denne typen, som de som er bindende til DNA, kan aktivere eller undertrykke gentranskripsjon, slik at de kalles henholdsvis "tvangsverdatorer" og "korrigert".
Regulering
Disse proteinene kan reguleres på to nivåer: i deres syntese og i deres aktivitet, som avhenger av forskjellige variabler og flere situasjoner.
Synteseforordning
Reguleringen av syntesen kan være relatert til uttrykket vevsspesifikk av visse transkripsjonsfaktorer. Et eksempel på dette kan være myodfaktoren, bare syntetisert i skjelettmuskelceller og er nødvendig for differensiering av dens udifferensierte fibroblaster.
Kan tjene deg: koblede generSelv om regulering av syntese er grunnleggende brukt for kontroll av genetisk uttrykk i spesifikke celletyper og vev, er dette ikke den eneste måten, siden syntese av faktorer som er involvert i induksjon av gener som deltar i responsen også er regulert til flere stimuli.
Aktivitetsregulering
En annen reguleringsmekanisme for transkripsjonsfaktorer er reguleringen av dens aktivitet, noe som har å gjøre med aktivering av andre eksisterende transkripsjonsfaktorer som utøver positive eller negative effekter på aktiviteten til en bestemt faktor.
Aktiveringen av disse "sekundære" faktorene skjer vanligvis gjennom forskjellige mekanismer som ligandforening, endringer i protein-protein, fosforyleringsinteraksjoner, blant andre.
Funksjoner og betydning
Transkripsjonsfaktorer deltar i en rekke prosesser som embryonal utvikling, vekst og differensiering, cellesykluskontroll, tilpasning til svingende miljøforhold, vedlikehold av spesifikke proteinsyntesemønstre av celler og vev, etc.
I planter har de for eksempel viktige funksjoner i forsvaret og som svar hendelser på forskjellige typer stress. Det er blitt bestemt at osteogenese hos dyr styres av transkripsjonsfaktorer, så vel som mange andre differensieringsprosesser av forskjellige cellelinjer.
Gitt viktigheten av disse proteinene i organismer, er det ikke uvanlig å tro at endringer i disse regulatoriske elementene vil forårsake alvorlige patologiske endringer.
Når det gjelder mennesker, kan patologier assosiert med transkripsjonsfaktorer være utviklingsforstyrrelser (på grunn av mutasjoner som gir inaktivering av transkripsjonsfaktorer, for eksempel), lidelser i hormonell respons eller kreftformer.
Referanser
- Alberts, f., Dennis, f., Hopkin, k., Johnson, a., Lewis, J., Raff, m.,... Walter, s. (2004). Essensiell cellebiologi. Abingdon: Garland Science, Taylor & Francis Group.
- Iwafuchi-doi, m., & Zaret, K. S. (2014). Pioneer transkripsjonsfaktorer i celleprogrammering. Gener og utvikling, 28, 2679-2692.
- Latchman, d. (1997). Transkripsjonsfaktorer: en oversikt. Int. J. Biochem. Celle. Biol., 29(12), 1305-1312.
- Latchman, d. S. (2007). Transkripsjonsfaktorer. Encyclopedia of Life Sciences, 1-5.
- Marie, s. J. (2008). Transkripsjonsfaktorer som kontrollerer osteoblastogenese. Arkiv for biokjemi og biofysikk, 473, 98-105.
- Pabo, c., & Sauer, r. T. (1992). Transkripsjonsfaktorer: Strukturelle familier og prinsipper for DNA -anerkjennelse. Annu. Rev., 61, 1053-1095.
- Singh, k. B., Foley, r. C., & Oñate-sánchez, l. (2002). Transkripsjonsfaktorer i planteforsvar og stressrespons. Nåværende mening i plantebiologi, 5, 430-436.
- « Bussegenskaper, mat, reproduksjon, habitat
- Klorofytter Karakteristikker, habitat, reproduksjon, mat »