

Mellomliggende filamenter struktur, typer, funksjoner
- 2587
- 37
- Anders Mathisen
De mellomfilamenter, Også kjent i litteratur som "ifs" (fra engelsk Mellomliggende filames), er en familie av uoppløselige cytosoliske fibrøse proteiner som er til stede i alle flercellede eukaryote celler.
De er en del av cytoskjelettet, som er et intracellulært filamentøst nettverk som hovedsakelig er ansvarlig for støtte fra cellestrukturen og forskjellige metabolske og fysiologiske prosesser som transport av vesikler, bevegelse og celleforskyvning, etc.
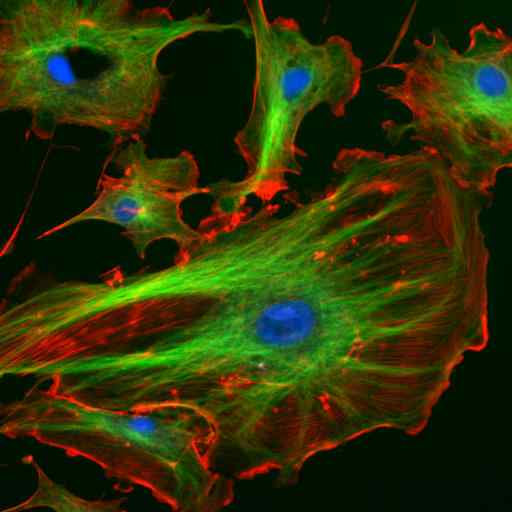
 Mikroskopi ved immunofluorescens av to proteiner av de mellomliggende filamentene til astrocytter (Vimentina og GFAP) (Kilde: Gerryshaw [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons)
Mikroskopi ved immunofluorescens av to proteiner av de mellomliggende filamentene til astrocytter (Vimentina og GFAP) (Kilde: Gerryshaw [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] via Wikimedia Commons) Sammen med mikrotubuli og mikrofilamenter deltar mellomfilamenter i den romlige organisasjonen av intracellulære organeller, i endocytose og eksocytoseprosesser og også i prosessene for celledeling og intercellulær kommunikasjon.
De første mellomliggende filamentene som ble studert og beskrevet var keratinene, en av de første typene proteiner hvis struktur ble analysert ved røntgendiffraksjon på 1930 -tallet.
Konseptet med mellomliggende filamenter ble imidlertid introdusert på 1980 -tallet av Lazarides, som beskrev dem som "mekaniske integratorer av cellulært rom", preget av deres uoppløselighet og deres evne til å gjense In vitro Etter denaturering.
Mange forfattere regnes som de "dempende" elementene i stress for dyreceller, siden de er mer fleksible filamenter enn mikrotubuli og mikrofilamenter. Ikke bare finnes de i cytoskjelettet, men de er også en del av nukleoesqueleto.
I motsetning til de andre fibrøse komponentene i cytoskjelettet, deltar ikke mellomliggende filamenter direkte i cellemobilitetsprosesser, men fungerer i strukturell vedlikehold og mekanisk resistens av celler.
[TOC]
Struktur
Kilde: http: // RSB.Info.NIH.GOV/IJ/IMAGER/[Public Domain] De mellomliggende filamentene har en omtrentlig diameter på 10 nm, strukturell karakteristikk som de ble navngitt, siden størrelsen er mellom størrelsene som tilsvarer myosin- og aktinfilamentene, som er henholdsvis rundt 25 nm og 7 nm.
De skiller seg strukturelt fra de to andre typene cytoskjelettfilamenter, som er kuleproteinpolymerer, der deres bestanddeler er forskjellige a-helikoidale fibrøse proteiner med stor lengde som er gruppert fra hverandre for å danne strukturer i form av tau eller tau.
Alle proteiner som utgjør mellomfilamentene har en lignende molekylær organisasjon, som består av et α-helicidal eller "tau" -domene som har forskjellige mengder "spoleformatorer" -segmenter av samme størrelse.
Dette spiralformede domenet blir flankert av et ikke-terminalt "hode" og en ikke-hypoidal "hale" i den C-terminale enden, og begge varierer både i størrelse og aminosyresekvens.
Innenfor sekvensen til disse to ytterpunktene er konsensusgrunner som er vanlige for de 6 typene kjente mellomfilamenter.
I virveldyr har "tau" -domenet til proteinet til de mellomliggende cytosoliske filamentene omtrent 310 aminosyreavfall, mens de cytosoliske proteiner fra virvelløse dyr og kjerneminatet har omtrent 350 aminosyrer i lengde.
Kan tjene deg: Reptiler: Hva er, egenskaper, klassifisering, reproduksjonmontering
Mellomliggende filamenter er "selvmåhet" -strukturer som ikke har enzymatisk aktivitet, som også skiller dem fra deres cytoskeletale kolleger (mikrotubuli og mikrofilamenter).
Disse strukturene er opprinnelig samlet som tetrammer av filamentøse proteiner som omfatter dem under påvirkning av bare monovalente kationer.
Disse tetrammerne er 62 nm lange, og deres monomerer er assosiert med hverandre sideveis for å danne "lengdeenheter" (UFL, av engelsk Enhetslegh-filmeringer), Det som er kjent som fase 1 av forsamlingen, som skjer veldig raskt.
UFL -ene er forløperne for de lange filamentene, og siden dimerer som består av dem binder hverandre i en antiparalle og forskjøvet måte, har disse enhetene et sentralt domene med to flankerende domener som fase 2 av forlengelsen skjer, hvor den langsgående union av andre UFL -er er gitt.
Under det som er blitt kalt som fase 3 av monteringen, oppstår den radielle komprimeringen av diameteren på filamentene, som produserer de modne mellomliggende filamentene på mer eller mindre 10 nm i diameter.
Funksjoner
Funksjonene til de mellomliggende filamentene avhenger betydelig av typen celle som blir vurdert, og i tilfelle av dyr (inkludert mennesker) er deres uttrykk regulert på en vevsspesifikk måte, så det avhenger også av typen vev enn under under studere.
Epitel, muskler, mesenkymale og glials og nevroner har forskjellige typer filamenter, spesialisert i henhold til funksjonen til cellene de tilhører.
Blant disse funksjonene er det viktigste strukturelt vedlikehold av celler og motstand mot forskjellig mekanisk stress, siden disse strukturene har en viss elastisitet som lar dem dempe forskjellige typer krefter pålagt celler.
Typer mellomliggende filamenter
Proteiner som utgjør mellomfilamenter tilhører en stor og heterogen familie av filamentøse proteiner som er kjemisk forskjellige, men som skilles ut i seks klasser i henhold til deres homologi av sekvenser (I, II, III, IV, V og VI).
Selv om det ikke er veldig vanlig, er forskjellige typer celler, under veldig spesielle forhold (utvikling, celletransformasjon, vekst, etc.) De kan co-express mer enn en klasse av mellomliggende filament som danner proteiner
Klasse I og II av mellomfilamenter: syre og grunnleggende keratiner
Keratinene representerer de fleste av proteinene fra mellomliggende filamenter, og hos mennesker representerer disse mer enn tre fjerdedeler av mellomfilamentene.
De har molekylvekter som varierer mellom 40 og 70 kDa og skiller seg fra andre mellomliggende glødetrådsproteiner på grunn av deres høye innhold i glycin og serin.
Kan tjene deg: Hedgehogs: Kjennetegn, habitat, reproduksjon, matDe er kjent som syre og basiske keratiner for sine isoelektriske punkter, som er mellom 4.9 og 5.4 for sure keratiner og mellom 6.1 og 7.8 for grunnleggende.
I disse to klassene er rundt 30 proteiner blitt beskrevet og er til stede spesielt i epitelceller, der begge typer protein "co-polimeriser" og danner sammensatte filamenter.
Mange av saken I keratiner av mellomliggende filamenter finnes i strukturer som hår, negler, horn, pigger og klør, mens klasse II er de mest tallrike i cytosol.
Klasse III av mellomliggende filamenter: Demine/vimentin -type proteiner
Fallene er et surt protein på 53 kDa som, avhengig av dens grad av fosforylering, har forskjellige varianter.
Noen forfattere har også kalt filamentene til å falle som "muskel mellomliggende filamenter", siden deres tilstedeværelse er ganske begrenset, selv om alle typer muskelceller i små mengder er.
I myofibriller finnes fossene i linje Z, så det antas at dette proteinet bidrar til de kontraktile funksjonene til muskelfibre når du jobber i foreningen av myofibrillaer og plasmamembran.
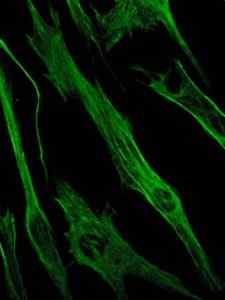 Fotografi av farging av vimentinproteinet, et protein av mellomliggende filamenter i epitel- og embryonale celler (kilde: Viktoriia Kosach [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Fotografi av farging av vimentinproteinet, et protein av mellomliggende filamenter i epitel- og embryonale celler (kilde: Viktoriia Kosach [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) På sin side er vimentin et protein som er til stede i mesenkymale celler. De mellomliggende filamentene dannet av dette proteinet er fleksible, og det er blitt bestemt at de motstår mange av de konformasjonsendringene som oppstår under cellesyklusen.
Det finnes i fibroblaster, i glatte muskelceller, i hvite blodlegemer og andre celler i dyresirkulasjonssystemet.
Klasse IV av mellomfilamenter: Neurofilamentøse proteiner
Også kjent som "neurofilamenter", inkluderer denne typen mellomfilamenter et av de grunnleggende strukturelle elementene i aksoner og neuronale dendritter; De er ofte assosiert med mikrotubulene som også utgjør disse strukturene.
Neurofilamenter av virveldyr er blitt isolert, og bestemmer at det er en 200, 150 og 68 kDa protein triplett som deltar i montering In vitro.
De skiller seg fra andre mellomliggende filamenter der de har sidearmer som "vedlegg" som er projisert fra periferien deres, og som fungerer i samspillet mellom nærliggende filamenter og andre strukturer.
Glialceller produserer en spesiell type mellomliggende filamenter kjent som glial mellomliggende filamenter, som strukturelt skiller.
Klasse V av mellomliggende filamenter: kjernefilamanter
Alle ark som er en del av nukleoesqueleto er faktisk mellomliggende filamentproteiner. Disse har mellom 60 og 75 kDa molekylvekt og finnes i kjernene til alle eukaryote celler.
Kan tjene deg: Trichuris TrichiuraDe er viktige for den interne organisasjonen av kjernefysiske regioner og for mange av funksjonene til denne essensielle organellen for eksistensen av eukaryoter.
Klasse VI av mellomfilamenter: Nestines
Denne typen mellomfilamenter veier mer eller mindre 200 kDa og er hovedsakelig i stamcellene i sentralnervesystemet. De kommer til uttrykk under neuronal utvikling.
Relaterte patologier
Det er flere sykdommer hos mennesker som er relatert til mellomfilamenter.
I noen typer kreft som ondartede melanomer eller brystkarsinomer, for eksempel, fører samekspresjonen av mellomliggende vimentin- og keratinfilamenter til differensiering eller interkonvertering av epitel- og mesenkymale celler.
Det er eksperimentelt demonstrert at dette fenomenet øker den trekkende og invasive aktiviteten til kreftceller, noe som har viktige implikasjoner i metastatiske prosesser som er karakteristiske for denne tilstanden.
Eriksson og samarbeidspartnere (2009) gjennomgår de forskjellige typer sykdommer og deres forhold til spesifikke mutasjoner i genene som er involvert i dannelsen av de seks typene mellomliggende filamenter.
Sykdommer relatert til kodende genmutasjoner for de to typene keratin er bullast epidermolyse, epidermolytisk hyperkeratose, hornhinnedystrofi, keratodermier og mange andre.
Type III mellomfilamenter er involvert i mange kardiomyopatier og forskjellige muskelsykdommer relatert hovedsakelig med dystrofier. I tillegg er de også ansvarlige for dominerende grå stær og noen typer sklerose.
Tallrike nevrologiske syndromer og lidelser er relatert til filamenter av type IV, for eksempel Parkinson Parkinson. Tilsvarende er genetiske defekter i filamenter av type V og VI ansvarlige for utvikling av forskjellige autosomale sykdommer og relatert til funksjonen til cellekjernen.
Eksempel på disse er Hutchinson-Gilford Progeria Syndrome, Emery-Dreifuss muskeldystrofi, blant andre.
Referanser
- Anderton, f. H. (1981). Mellomfilamler: En familie av homologe strukturer. Journal of Muscle Research and Cell Motilitet, 2(2), 141-166.
- Eriksson, J. OG., Pallari, h., Robert, d., Eriksson, J. OG., Dechat, t., Glis, f.,... Goldman, r. D. (2009). Innføring av mellomliggende filamorer: Fra oppdagelse til sykdom. Journal of Clinical Investigation, 119(7), 1763-1771.
- Fuchs, e., & Weber, K. (1994). Mellomfilamler: struktur, dynamikk, funksjon og sykdom. Annu. Rev. Biochem., 63, 345-382.
- Hendrix, m. J. C., Seftor, e. TIL., Chu, og. W., Trevor, k. T., & Seftor, r. OG. B. (nitten nittiseks). Rollen til mellomliggende fillamenter i migrasjon, invasjon og metastase. Kreft og metastaseanmeldelser, femten(4), 507-525.
- Herrmann, h., & Aebi, u. (2004). Mellomliggende filn: Molekylær struktur, monteringsmekanisme og integrasjon i funksjonelt intracellulære stillaser distinkt. Årlig gjennomgang av biokjemi, 73(1), 749-789.
- Herrmann, h., & Aebi, u. (2016). Mellomfilamler: struktur og montering. Kald vårhavnperspektiver i biologi, 8, 1-22.
- McLean, i., & Lane, B. (nitten nitti fem). Mellomliggende filamorer i Diew. Nåværende mening i cellebiologi, 7(1), 118-125.
- Steinert, p., & Roop, d. (1988). Molekylær og cellulær biologi av mellomfilmer. Årlig gjennomgang av biokjemi, 57(1), 593-625.
- Steinert, p., Jones, J., & Goldman, r. (1984). Mellomliggende filames. Journal of Cell Biology, 99(1), 1-6.
- « Gastrotriske egenskaper, habitat, reproduksjon, mat
- Myke moskeegenskaper, taksonomi, habitat, bruker »