

Oksidative fosforyleringsstadier, funksjoner og hemmere
- 3914
- 1263
- Magnus Sander Berntsen
De oksidativ fosforylering Det er en prosess der ATP -molekyler blir syntetisert fra ADP og PYo (Uorganisk fosfat). Denne mekanismen utføres av bakterier og eukaryote celler. I eukaryote celler utføres fosforylering i mitokondriell matrise av ikke -fotosyntetiske celler.
ATP -produksjon er regissert ved overføring av elektroner fra NADH eller FADH Coenzymes2 til o2. Denne prosessen representerer den største energiproduksjonen i cellen og er avledet fra nedbrytningen av karbohydrater og fett.
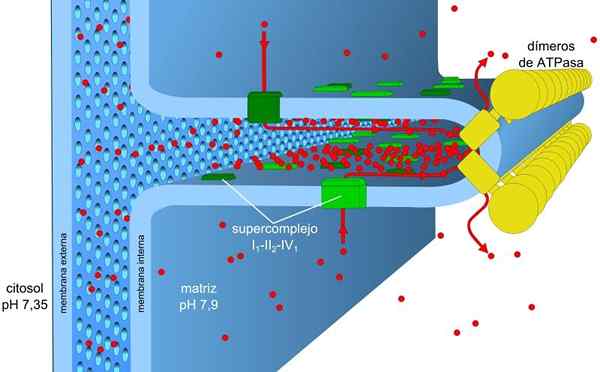
Kilde: Robot8a [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] Energien som er lagret i belastnings- og pH -gradienter, også kjent som protonisk motorisk kraft, gjør at denne prosessen kan utføres. Protongradienten som genereres, får den ytre delen av membranen til å ha en positiv belastning på grunn av konsentrasjonen av protoner (H+) og mitokondriell matrise er negativ.
[TOC]
Der oksidativ fosforylering skjer?
Elektrontransport og oksidative fosforyleringsprosesser er assosiert med en membran. I prokaryoter blir disse mekanismene utført gjennom plasmamembranen. I eukaryote celler er de assosiert med mitokondriemembranen.
Antall mitokondrier som finnes i cellene varierer i henhold til celletypen. For eksempel, hos pattedyr, mangler erytrocytter disse organellene, mens andre celletyper, for eksempel muskelceller, kan ha millioner av dem.
Den mitokondrielle membranen består av en enkel ytre membran, en noe mer kompleks indre membran, og midt i dem er det intermembranale rommet, der mange enzymer avhengig av ATP er lokalisert.
Den ytre membranen inneholder et protein kalt porina som danner kanalene for enkel diffusjon av små molekyler. Denne membranen er ansvarlig for å opprettholde strukturen og formen på mitokondriene.
Den indre membranen har større tetthet og er rik på protein. Det er også vanntett for molekyler og ioner, så for å krysse det, trenger de intermembranale proteiner som transporterer dem.
Innenfor matrisen strekker foldene av den indre membranen seg, og danner rygger som lar den ha et stort område i et lite volum.
Cell Energy Central
Mitokondriene regnes som det cellulære energisenteret. I den er enzymene som er involvert i prosessene i cytrinsyresyklusen, oksidasjon av fettsyrer og redoksenzymer og proteiner av elektrontransport og PhD -fosforylering.
Konsentrasjonsgradienten av protonene (pH -gradient) og gradienten av belastning eller elektrisk potensial i den indre membranen til mitokondriene, er årsaken til den praktoniske motoriske kraften. Den lille permeabiliteten til den indre membranen for ioner (annet enn h+) lar mitokondriene ha en stabil spenningsgradient.
Elektronisk transport, protoner pumping og ATP oppnår samtidig i mitokondriene, takket være den protoniske motoriske styrken. PH -gradienten opprettholder sure forhold under intermembran og mitokondriell matrise med alkaliske forhold.
For hver to elektroner overført til O2 Cirka 10 protoner pumpes av membranen, og skaper en elektrokjemisk gradient. Energien løsrevet i denne prosessen produseres gradvis ved å føre elektroner gjennom transportørkjeden.
Stadier
Energien som frigjøres under oksydreduksjonsreaksjoner av NADH og FADH2 Det er betydelig høyt (ca. 53 kcal/mol for hvert par elektroner), så for å brukes i fremstilling av ATP -molekyler, må den gradvis produseres med passering av elektroner gjennom transportører.
Disse er organisert i fire komplekser som ligger i den indre mitokondrielle membranen. Koblingen av disse reaksjonene på ATP -syntese utføres i et femte kompleks.
Kan tjene deg: B -lymfocytter: Kjennetegn, struktur, funksjoner, typerElektrontransportkjede
NADH overfører et par elektroner som kommer inn i komplekset i elektrontransportkjeden. Elektronene blir overført til mononukleotidet i flavina, og deretter til ubiquinon (koenzym Q) gjennom en jernsugar-transportør. Denne prosessen frigjør en stor mengde energi (16,6 kcal/mol).
Ubiquinona transporterer elektroner gjennom membranen til kompleks III. I dette komplekset går elektronene gjennom cytokromer B og C1 Takk til en jern-sukker-transportør.
Fra kompleks III passerer elektroner til IV -komplekset (cytokrom c oksidase), overført en av cytokrom c (perifert membranprotein). I IV -komplekset passerer elektronene gjennom et par kobberioner (Cutil2+), deretter til cytokrom ctil, Så til et annet par kobberioner (Cub2+) og fra dette til cytokrom til3.
Endelig overføres elektroner til O2 som er den siste akseptoren og danner et vannmolekyl (h2O) For hvert mottatt elektroner. Passering av elektroner fra kompleks IV til O2 Det genererer også mye gratis energi (25,8 kcal/mol).
Succinato CoQ -reduktase
II -komplekset (CoQ -reduktase succinat) mottar et par elektroner fra sitronsyresyklusen, på grunn av oksidasjon av et succinatmolekyl til fumarat. Disse elektronene blir overført til kjepphesting, og passerer deretter gjennom en jern-azufre-gruppe, til Ubiquinona. Fra dette koenzymet går de til Complex III og følger ruten beskrevet tidligere.
Energien som frigjøres i elektronoverføringsreaksjonen på kjepphest er ikke nok til å øke protonene gjennom membranen, slik at det i dette trinnet i kjeden ikke er noen praktonisk motorisk kraft, og følgelig gir FADH mindre H+ enn nadh.
Energikobling eller transduksjon
Energien som genereres i elektrontransportprosessen som tidligere er beskrevet, skal brukes til ATP -produksjon, reaksjon katalysert av ATP -syntasen eller kompleks enzym V. Bevaring av slik energi er kjent som energikobling, og mekanismen har vært vanskelig å karakterisere.
Flere hypoteser er beskrevet for å beskrive denne energitransduksjonen. Den best aksepterte er den kjemosmotiske koblingshypotesen, beskrevet nedenfor.
Kjemosmotisk kobling
Denne mekanismen foreslår at energien som brukes til ATP -syntese kommer fra en protonisk gradient i cellemembraner. Denne prosessen griper inn i mitokondrier, kloroplaster og bakterier og er knyttet til elektrontransport.
I- og IV -kompleksene av elektronisk transport fungerer som protonpumper. Disse lider av konformasjonsendringer som lar dem pumpe protonene til det intermembranale rommet. I IV -komplekset for hvert par elektroner pumpes to protoner utenfor membranen og to til gjenstår i matrisen som danner H2ENTEN.
Ubiquinona i Complex III aksepterer protoner av komplekser I og II og frigjør dem utenfor membranen. I og III -komplekser tillater hver passering av fire protoner for hvert par transporterte elektroner.
Den mitokondrielle matrisen har lav konsentrasjon av protoner og negativt elektrisk potensial mens det intermembranale rommet presenterer de inverse forholdene. Strømmen av protoner gjennom denne membranen innebærer den elektrokjemiske gradienten som lagrer den nødvendige energien (± 5 kcal/mol av proton) for ATP -syntese.
ATP -syntese
ATP -syntetaseenzymet er det femte komplekset som er involvert i oksidativ fosforylering. Er ansvarlig for å dra nytte av energien til den elektrokjemiske gradienten for å danne ATP.
Dette transmembraale proteinet består av to komponenter: f0 og f1. Komponenten f0 tillater protons retur til mitokondriell matrise som fungerer som en kanal og f1 katalyserer syntesen av ATP via ADP og PYo, Bruke energien til nevnte retur.
Kan tjene deg: membran transportørproteiner: funksjoner og typerATP -synteseprosessen krever en strukturell endring i f1 og montering av komponenter f0 og f1. Translokasjon av protoner gjennom f0 Det forårsaker konformasjonsendringer i tre undermunner F1, slik at dette kan fungere som en rotasjonsmotor, og styrer dannelsen av ATP.
Underenheten som er ansvarlig for foreningen av ADP med PYo Går fra en svak tilstand (L) til en aktiv (t). Når ATP dannes, passerer en andre underenhet til en åpen tilstand (O) som tillater frigjøring av dette molekylet. Etter at ATP er utgitt, går denne underenheten fra åpen tilstand til en inaktiv tilstand (L).
ADP- og P -molekyleneYo De blir med i en underenhet som har gått fra en stat eller staten l.
Produkter
Elektron transportørkjeden og fosforylering produserer ATP -molekyler. NADH -oksidasjon produserer omtrent 52,12 kcal/mol (218 kJ/mol) av fri energi.
Den globale reaksjonen for NADH -oksidasjon er:
Nadh+1⁄2 eller2 +H+ ↔ H2O+nad+
Overføringen av elektroner fra Nadh og Fadh2 Det skjer gjennom flere komplekser, slik at endring av fri energi Δg ° kan deles inn i mindre “pakker”, som er koblet til ATP -syntese.
Oksidasjonen av et NADH -molekyl genererer syntesen av tre ATP -molekyler. Mens oksidasjonen av et FADH -molekyl2 Det er knyttet til syntesen av to ATP.
Disse koenzymene kommer fra prosessene med glykolyse og sitronsyresyklus. For hvert nedbrutt glukosemolekyl er 36 eller 38 ATP -molekyler ender, avhengig av cellens plassering. I hjernen og skjelettmuskel 36 produseres ATP mens 38 ATP produseres i muskelvev.
Funksjoner
Alle organismer, encellede og flercellede, trenger en minimumsenergi i cellene sine for å utføre prosessene i dem, og opprettholder på sin side viktige funksjoner i den komplette kroppen.
Metabolske prosesser krever at energi skal utføres. Det meste av den brukbare energien oppnås ved nedbrytning av karbohydrater og fett. Denne energien stammer fra den oksidative fosforyleringsprosessen.
Oksidativ fosforyleringskontroll
ATP -brukshastigheten i cellene, kontrollerer syntesen av det samme, og på sin side, på grunn av koblingen av oksidativ fosforylering med elektrontransportkjeden, regulerer den også den elektroniske transporthastigheten.
Oksidativ fosforylering har en streng kontroll som sikrer at ATP ikke genereres raskere enn den konsumeres. Det er visse trinn i elektrontransportprosessen og den koblede fosforylering som regulerer energiproduksjonshastigheten.
Koordinert kontroll av ATP -produksjonen
De viktigste energiproduksjonsveiene (celle ATP) er glykolyse, syklusen av sitronsyre og oksidativ fosforylering. Koordinert kontroll av disse tre prosessene regulerer ATP -syntese.
Fosforyleringskontroll av ATP -massehandlingsforholdet avhenger av det nøyaktige bidraget til elektroner i transportørkjeden. Dette avhenger igjen av forholdet [NADH]/[NAD+] som er bevart forhøyet av virkningen av glykolyse og sitronsyresyklusen.
Denne koordinerte kontrollen utføres ved å regulere glykolysekontrollpunkter (Citrate hemmet PFK) og cytricyre-syklusen (dehydrogenase-pyruvat, cutease citrat, kløende dehydrogenase og a-zetoglutaratdehydrogenase).
Kontroll av akseptor
IV -komplekset (cytokrom c oksidase) er et enzym regulert av et av dets underlag, det vil si at aktiviteten styres av den reduserte cytokrom c (C2+), som igjen er i balanse med forholdet mellom konsentrasjonene mellom [NADH]/[NAD+] og masseaksjonsforholdet til [ATP]/[ADP] + [PYo].
Det kan tjene deg: Tone: Kjennetegn og funksjonerJo høyere [nadh]/[nad+] og senk [ATP]/[ADP]+[PYo], mer konsentrasjon vil være cytokrom [c2+] og større vil være aktiviteten til kompleks IV. Dette tolkes for eksempel hvis vi sammenligner organismer med forskjellige hvileaktiviteter og høy aktivitet.
I et individ med høy fysisk aktivitet, forbruk av ATP og derfor hydrolyse til ADP + PYo Det vil være veldig høyt, og generere en forskjell i masseaksjonsforholdet som forårsaker en økning i [C2+og derfor en økning i ATP -syntese. I et hvilende individ oppstår den omvendte situasjonen.
Til slutt øker hastigheten på oksidativ fosforylering med konsentrasjonen av ADP i mitokondriene. Denne konsentrasjonen avhenger av ADP-ATP-translokatorer med ansvar for transport av adeninnukleotider og pYo Fra cytosol til mitokondriell matrise.
DECACOPLANTS AGENTER
Oksidativ fosforylering påvirkes av visse kjemiske midler, som lar elektronisk transport fortsette uten fosforylering av ADP, avkoble til produksjon og bevaring av energi.
Disse midlene stimulerer mitokondriene oksygenforbrukshastighet i fravær av ADP, og forårsaker også en økning i ATP -hydrolyse. De virker ved å eliminere en mellomledd eller bryte en energitilstand fra elektrontransportørkjeden.
2,4-dinitrofenol, en svak syre som passerer gjennom mitokondrielle membraner, er ansvarlig for å spre protonsgradienten, fordi de blir med dem på den sure siden og slipper dem på den grunnleggende siden.
Denne forbindelsen ble brukt som en "slankende pille" fordi den ble funnet at den ga en økning i pusten, derfor en økning i metabolskhastigheten og tilhørende vekttap. Det ble imidlertid vist at dens negative effekt til og med kunne forårsake død.
Spredningen av den protoniske gradienten produserer varme. Brun fettvevsceller bruker avkobling, hormonelt kontrollert, for å produsere varme. Dvalemodus og nyfødte som mangler hår består av dette vevet som tjener dem som et slags termisk teppe.
Hemmere
Hemmende forbindelser eller midler forhindrer både forbruk av eller2 (elektronisk transport) som tilhørende oksidativ fosforylering. Disse midlene unngår dannelse av ATP gjennom bruk av energi produsert i elektronisk transport. Derfor stopper transportkjeden da slikt energiforbruk ikke er ordnet.
Oligomicin antibiotika fungerer som en fosforyleringsinhibitor i mange bakterier, og forhindrer ADP -stimulering fra ATP -syntese.
Det er også ionophorøse midler, som utgjør fett -oppløselige komplekser med kationer som k+ og na+, og passere gjennom mitokondriell membran med disse kationene. Mitokondriene bruker deretter energi produsert i elektronisk transport for å pumpe kationer i stedet for å syntetisere ATP.
Referanser
- Alberts, f., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, P. (2004). Essensiell cellebiologi. New York: Garland Science.
- Cooper, g. M., Hausman, r. OG. & Wright, n. (2010). Cellen. (s. 397-402). Marbán.
- Devlin, t. M. (1992). Biokjemiets lærebok: med kliniske korrelasjoner. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, C. M. (2008). Biokjemi. Thomson Brooks/Cole.
- Lodish, h., Darnell, J. OG., Berk, a., Kaiser, ca. TIL., Krieger, m., Scott, m. P., & Matsudaira, P. (2008). Mollekylær cellebiologi. Macmillan.
- Nelson, d. L., & Cox, m. M. (2006). Lehninger prinsipper for biokjemi 4. utgave. Ed Omega. Barcelona.
- Voet, d., & Voet, j. G. (2006). Biokjemi. Ed. Pan -American Medical.
- « Euglenophyta -egenskaper, reproduksjon, ernæring, klassifisering
- Glukosider trening, funksjon og typer/grupper »