

Glyceraldehyd 3-fosfat (G3P) struktur, funksjoner
- 5058
- 365
- Theodor Anders Hopland
Han Glyceraldehyd 3-fosfat (GAP) Det er en metabolitt av glykolyse (navnet kommer fra det greske; glykos = søt eller sukker; lysis = brudd), som er en metabolsk vei som konverterer glukosemolekylet til to pyruvatmolekyler for å produsere energi i form av adenosinprøfat (ATP ).
I celler kobler glyceraldehyd 3-fosfat glykolyse med glukoneogenese og pentosefosfatbanen. I fotosyntetiske organismer brukes glyseraldehyd 3-fosfat, fra karbondioksidfiksing, til sukkerbiosyntese. I leveren produserer fruktosemetabolismen gap, som er integrert i glykolyse.
 Kilde: Benjah-BMM27 [Public Domain]
Kilde: Benjah-BMM27 [Public Domain] [TOC]
Struktur
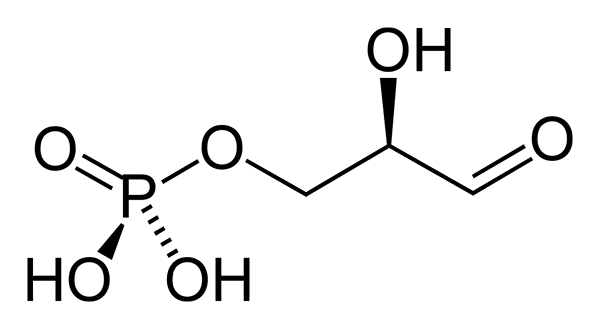
3-fosfatglyseraldehydet er et fosforylert sukker som har tre karbonatomer. Dens empiriske formel er c3H7ENTEN6P. Aldehydgruppen (-CHO) er karbon 1 (C-1), hydroksymetylengruppen (-CHAH) er karbon 2 (C-2) og hydroksymetylgruppen (-CH2Å) er karbon 3 (C3). Denne siste danner en kobling med fosfatgruppen (Fosfoester Link).
3-fosfatglyseraldehydkonfigurasjonen i C-2 quiral er d. Ved konvensjon, med hensyn til det quirale karbonet, i projeksjonen av Fischer, er aldehydgruppen representert oppover, hydroksymetyl-fosfatgruppen ned, hydroksylgruppen til høyre og hydrogenatom til venstre.
Kjennetegn
3-fosfatglyseraldehydet har en molekylmasse 170,06 g/mol. Standard Gibbs Free Energy Change (Δgº) for enhver reaksjon må beregnes ved å tilsette variasjonen av produktenees frie energi, og trekke fra summen av variasjonen av reaktantens frie energi.
Det kan tjene deg: Metazoa: Kjennetegn, typer, habitat og sykdommerPå denne måten bestemmes fri energivariasjon (Δgº) av glyseraldehyd 3 -fosfat, som er -1,285 kJ × mol-1. Etter konvensjon, i standardtilstanden 25 ºC og 1 atm, er den frie elementene frie energi null.
Funksjoner
Glykolyse og glukoneogenese
Glykolyse er til stede i alle celler. Det er delt inn i to faser: 1) energiinvesteringsstadium og metabolitter syntese med høyt fosfatgruppeoverføringspotensial, for eksempel glyseraldehyd 3-fosfat (GAP); 2) ATP -syntesefase fra molekyler med høyt overføringspotensial for fosfatgruppe.
3-fosfatglyseraldehyd og dihydroksyacetonfosfat. 3-fosfatglyseraldehyd blir omgjort til 1,3-bifosfoglyserat (1,3 bpg), ved hjelp av en reaksjon katalysert av enzymet gap dehydrogenase.
Dehydrogenase -gapet katalyserer oksidasjonen av aldehydkarbonatom og overfører en fosfatgruppe. Dermed dannes en blandet anhydrid (1,3 kmpg) der syregruppen, og fosforatomet er utsatt for den nukleofile angrepsreaksjonen.
Deretter, i en reaksjon katalysert av 3-fosfoglyseratkinasen, overfører 1,3 bpg fosfatgruppen fra karbon 1 til ADP, og danner ATP.
Fordi reaksjonene katalysert av aldolasen, er dehydrogenase-gapet og 3-fosfoglyseratkinasen i likevekten (Δgº ~ 0), er derfor en del av glukoneogenesens vei (eller den nye syntesen av glukose).
Vía de la Pentosa fosfat og Calvin Cycle
I pentosefosfatveien dannes 3-fosfat (GAP) glyseraldehyd og 6-fosfat fruktose (F6P) ved skjæring og dannelsesreaksjoner av C-C-bindinger, fra pentose, xylulose 5-fosfat og ribose 5-fosfat.
Kan tjene deg: Hyracotherium: Kjennetegn, ernæring, arter, reproduksjon3-fosfat glyseraldehyd kan følge banen til glukoneogenese og form 6-fosfat glukose, som fortsetter banen til pentosefosfatet. Glukose kan oksideres fullstendig ved å produsere seks co -molekyler2 Gjennom det oksidative stadiet av Pontosa Fosfatvei.
I Calvin -syklusen, CO2 Det er satt som 3-fosfoglyserat, i en reaksjon katalysert av den ribuløse bifosfatkarboksylase. Deretter reduseres 3-fosfoglyserat med NADH ved handling av et enzym kalt gap dehydrogenase.
2 gapmolekyler for biosyntese av en heksose, for eksempel glukose, som tjener for stivelse eller cellulosebiosyntese i planter i planter er nødvendig.
Fruktosemetabolisme
Fruktokinase-enzym katalyserer fosforylering av fruktose av ATP i C-1, og danner 1-fosfat fruktose. Aldolase A, som finnes i muskelen, er spesifikk for fruktose 1,6-bifosfat som et underlag. Aldolase B finnes i leveren og er spesifikk for 1-fosfat fruktose som et underlag.
B-aldolasen katalyserer det aldoliske bruddet av fruktose 1-fosfat og produserer dihydroksyacetonfosfat og glyseraldehyd. Glyseraldehydkinasen katalyserer fosforylering av glyseraldehyd gjennom ATP, og danner en glykolytisk mellomledd, 3-fosfatglyseraldehyd (GAP).
I en annen rute blir glyseraldehyd transformert til glyserol av en alkoholdehydrogenase som NADH bruker som et elektrondonorsubstrat. Deretter, glyserol fosforyler kinase glyserol av ATP, danner glyserol fosfat. Denne siste metabolitten er reoxy, og danner dihydroksyacetonfosfat (DHAP) og NADH.
DHAP konverteres til gap av de tre isfosfatenzymene. På denne måten konverteres fruktose til glykolysemetabolitter. Imidlertid kan fruktose levert intravenøst forårsake alvorlig skade, som består av den drastiske reduksjonen i intracellulært fosfat og ATP. Til og med melkesyreoser oppstår.
Det kan tjene deg: Chihuahua Flora og Fauna: Enestående arterFruktoseskader skyldes at det ikke har reguleringspunktene som glukosekatabolisme normalt har. Først kommer fruktose inn i musklene gjennom GLUT5, som er uavhengig av insulin.
For det andre konverteres fruktose direkte til gap, og på denne måten går den ikke gjennom reguleringen av fosfofutokinase (PFK) enzym i begynnelsen av glykolyse.
Via Entner-Doudoroff
Glykolyse er den universelle ruten for glukosekatabolisme. Noen bakterier bruker imidlertid ENTNER-DOUDOROFF-veien. Denne ruten innebærer seks trinn katalysert av enzymer, der glukose blir forvandlet til gap og pyruvato, som er to sluttprodukter av denne veien.
Gap og pyruvat blir transformert til etanol ved alkoholfermenteringsreaksjoner.
Referanser
- Berg, J. M., Tymoczco, J. L., STRYER, L. 2015. Biokjemi. Til kort kurs. W. H. Freeman, New York.
- Miesfeld, r. L., McEvoy, m. M. 2017. Biokjemi. W. W. Norton, New York.
- Nelson, d. L., Cox, m. M. 2017. Lehninger prinsipper for biokjemi. W. H. Freeman, New York.
- Saway J. G. 2004. Metabolisme på et øyeblikk. Blackwell, Malden.
- Voet, d., Voet, J. G., Pratt, c. W. 2008. Grunnleggende om biokjemi: Livet på molekylært nivå. Wiley, Hoboken.