

Mitosefaser, egenskaper, funksjoner og organismer
- 4023
- 634
- Dr. Andreas Hopland
De mitose Det er en celledelingsprosess, der en celle produserer genetisk identiske datterceller; For hver celle genereres to "døtre" med samme kromosombelastning. Denne inndelingen utføres i de somatiske cellene fra eukaryote organismer.
Denne prosessen er et av stadiene i cellesyklusen til eukaryote organismer, som er forstått i 4 faser: S (DNA -syntese), M (celledeling), G1 og G2 (mellomfaser der mRNA og proteiner forekommer). Sammen regnes faser G1, G2 og S som et grensesnitt. Den nukleære og cytoplasmatiske inndelingen (mitose og cytokinesis) utgjør det siste trinnet i cellesyklusen.
MYDOSIS Oversikt. Kilde: ViewAprabha [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] På molekylært nivå initieres myitose ved aktivering av en kinase (protein) kalt MPF (modningsfremmende faktor) og den påfølgende fosforylering av et betydelig antall cellekomponenterkomponenter. Sistnevnte lar cellen presentere de morfologiske endringene som er nødvendige for å utføre delingsprosessen.
Mitose er en aseksuell prosess, siden stamfadercellen og dens døtre har nøyaktig den samme genetiske informasjonen. Disse cellene er kjent som diploid ved å bære den komplette kromosombelastningen (2N).
Meiose er derimot prosessen med celledeling som resulterer i seksuell reproduksjon. I denne prosessen repliserer en diploid stamcelle dets kromosomer og deler deretter to ganger på rad (uten å replikere dens genetiske informasjon). Til slutt genereres 4 datterceller med bare halvparten av kromosombelastningen, som kalles haploider (n).
[TOC]
Generaliteter av mitose
Mitose i encellede organismer produserer vanligvis datterceller veldig lik foreldrene. I motsetning til dette, under utviklingen av flercellede vesener, kan denne prosessen forårsake to celler med noen forskjellige egenskaper (til tross for at de er genetisk identisk).
Denne celledifferensieringen gir opphav til de forskjellige celletypene som utgjør de flercellede organismer.
I løpet av en organismes levetid forekommer cellesyklusen kontinuerlig, og danner stadig nye celler som igjen vokser og forbereder seg på å dele seg gjennom myitose.
Cellulær vekst og inndeling er regulert av mekanismer, for eksempel apoptose (programmert celledød), som tillater å opprettholde en balanse, og unngå overflødig vekst av vev. På denne måten sikres det at mangelfulle celler erstattes av nye celler, i henhold til organismens krav og behov.
Hvilken relevans har denne prosessen?
Evnen til å reprodusere er en av de viktigste egenskapene til alle organismer (fra encellulær til flercellulær) og cellene som komponerer den. Denne kvaliteten gjør det mulig å sikre kontinuiteten i din genetiske informasjon.
Forståelsen av prosessene med mitose og meiose, har hatt en grunnleggende rolle i å forstå de spennende celleegenskapene til organismer. For eksempel, egenskapen til å holde antall kromosomer fra en celle til en annen i et individ, og mellom individer av samme art.
Når vi lider av en slags kutt eller sår i huden vår, observerer vi hvordan den skadede huden blir utvunnet i noen dager. Dette skjer takket være myitoseprosessen.
Faser og dens egenskaper
Generelt følger mytosen den samme prosesssekvensen (faser) i alle eukaryote celler. I disse fasene skjer det mange morfologiske forandringer i cellen. Blant dem kondensasjon av kromosomene, brudd på kjernefysisk membran, separasjon av cellen fra den ekstracellulære matrisen og andre celler, og inndelingen av cytoplasmen.
I noen tilfeller blir kjernefysisk inndeling og cytoplasmatisk inndeling betraktet som forskjellige faser (henholdsvis mitose og cytokinesis).
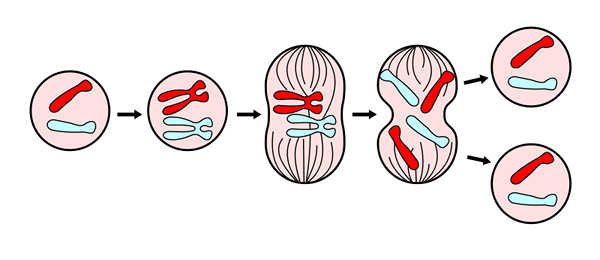
For en bedre studie og forståelse av prosessen er seks (6) faser blitt utpekt, kalt: Propase, lovet, metafase, anafase og telofase, og deretter vurderer cytokinesis som en sjette fase, som begynner å utvikle seg under anafase.
 Telophase er den siste fasen av mitose. Hentet fra https: // commons.Wikimedia.Org/wiki/fil: mitosepanel.Jpg. Via Wikimedia Commons
Telophase er den siste fasen av mitose. Hentet fra https: // commons.Wikimedia.Org/wiki/fil: mitosepanel.Jpg. Via Wikimedia Commons Disse fasene har blitt studert siden det nittende århundre gjennom lysmikroskopet, så i dag er de lett gjenkjennelige i henhold til de morfologiske egenskapene til cellen, så som kromosomal kondensasjon, og dannelsen av den mitotiske spindelen.
Profase
 Profase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons
Profase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons Profase er den første synlige manifestasjonen av celledelingen. I denne fasen kan du se utseendet til kromosomer som skillelige former, på grunn av den progressive komprimeringen av kromatin. Denne kondensasjonen av kromosomer begynner med fosforylering av Histona H1 -molekyler av MPF -kinasen.
Kondensasjonsprosessen består av sammentrekning og derfor reduksjon av størrelsen på kromosomene. Dette skjer på grunn av rulling av kromatinfibre, og produserer lettere displarbare strukturer (mitotiske kromosomer).
Kromosomene som tidligere er duplisert i perioden av cellesyklusen, skaffer seg et dobbelt glødetråd, kalt søsterkromatider, disse filamentene forblir samlet gjennom en region som kalles Centromero. I denne fasen forsvinner også nukleolien.
Det kan tjene deg: cromafinceller: egenskaper, histologi, funksjonerMitotisk spindeldannelse
Av Silvia3 [GFDL (http: // www.gnu.Org/copyleft/fdl.HTML) eller CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons Under profasen dannes den mitotiske spindelen, bestående av mikrotubuli og proteiner som utgjør et sett med fibre.
Når spindelen dannes, blir mikrotubulene i cytoskjelettet (ved deaktivering av proteinene som opprettholder strukturen) deassesemblet, noe som gir nødvendig materiale for dannelse av nevnte mitotisk spindel.
Sentrene (en organell uten membran, funksjonell i cellesyklusen), duplikat i grensesnittet, fungerer som monteringsenheten til spindelens mikrotubuli. I dyreceller har sentrering i sentrum, et par sentrioler; Men disse er fraværende i de fleste planteceller.
Dupliserte sentre, de begynner å skille den ene fra den andre mens spindelens mikrotubuli er samlet i hver av dem, og begynner å migrere til de motsatte ender av cellen.
På slutten av profasen begynner bruddet på atominnpakningen, som forekommer i separate prosesser: MAS -avgangen til kjernefysisk pore, atomarket og kjernefysiske membraner. Denne pausen lar mitotisk spindel og kromosomer begynne å samhandle.
Love
Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] På dette stadiet har kjernekonvolutten blitt helt fragmentert, så spindelens mikrotubuli invaderer dette området, og samhandler med kromosomer. De to sentrene har skilt seg, hver i polene til den mitotiske spindelen, i motsatte ytterpunkter av cellene.
Nå inkluderer den mitotiske spindelen mikrotubulene (som strekker seg fra hver sentrering til sentrum av cellen), sentrene og et par osterer (strukturer med radiell distribusjon av korte mikrotubuli, som er distribuert fra hver sentrering).
Kromatidene utviklet hver, en spesialisert proteinstruktur, kalt Cinetocoro, lokalisert i sentromere. Disse knetokoroene er plassert i motsatte retninger, og noen mikrotubuli festes, kalt mikrotubuli av Cinnetocoro.
Disse mikrotubulene festet til Cinetocoro begynner å bevege kromosomet fra slutten de utvider; Noen fra en stolpe og andre av motsatt pol. Dette skaper en "trekk- og krymp" -effekt som når de stabiliseres, lar kromosomet ende mellom endene av cellen.
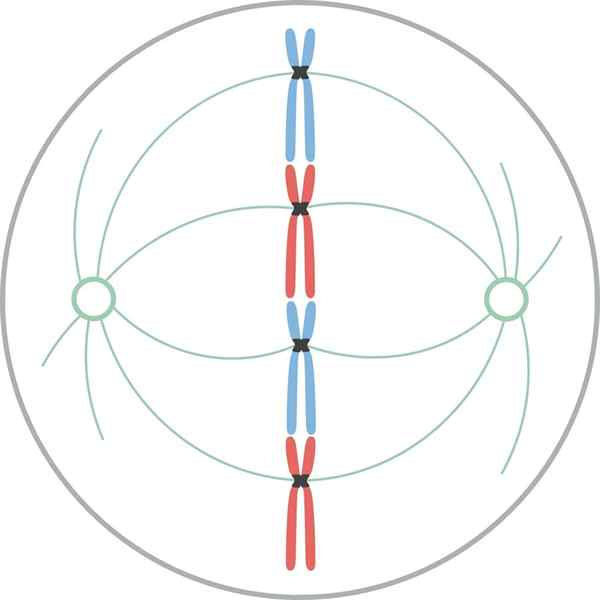
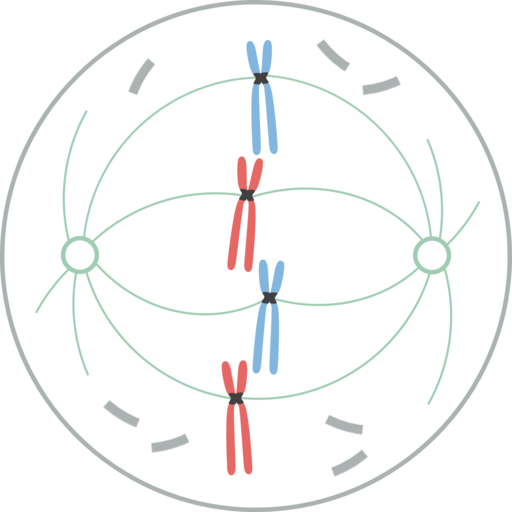
Metafase
Kromosomer justert i ekvatorialplaten til cellen under mytisk metafase I metafase er sentrene lokalisert i motsatte ender av cellene. Spindelen viser en klar struktur, i hvis sentrum kromosomene er lokalisert. Sentromerene til disse kromosomene er festet til fibrene og justert i et tenkt plan kalt metafasisk plakett.
Kromatid cipnetocoros forblir festet til mikrotubuli av cinetocoro. Mikrotubuli som ikke holder seg til Ketocoros og strekker seg fra motsatte poler av spindelen, samhandler nå med hverandre. På dette tidspunktet er mikrotubulene fra Osteres i kontakt med plasmamembranen.
Denne veksten og interaksjonen mellom mikrotubuli fullfører strukturen til den mitotiske spindelen, og gir et utseende "fuglebur".
Morfologisk er denne fasen den som virker mindre endringer, så den ble betraktet som en hvilefase. Selv om de ikke er lett på nytt, oppstår imidlertid mange viktige prosesser i det, i tillegg til å være det lengste stadiet av mitose.
Anafase
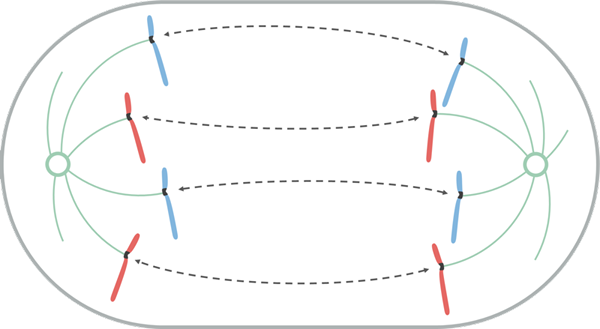 Kilde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons
Kilde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons Under anafasen begynner hvert par kromatider å skille seg (ved inaktivering av proteinene som holder dem sammen). Separate kromosomer beveger seg mot motsatte ender av cellen.
Denne migrasjonsbevegelsen skyldes det faktum at mikrotubulene i Catocoro de acortan, genererer en "trekk" -effekt som gjør hvert kromosombevegelse, fra sin centromero. Avhengig av plasseringen av sentromeren i kromosomet, kan det ta under forskyvningen en bestemt form som V eller J.
Mikrotubulene holdt seg ikke til Knetocoro, vokser og forlenges ved tubulinadhesjon (protein) og ved virkningen av motoriske proteiner som beveger seg på dem, slik at kontakten mellom dem kan stoppe. Når de beveger seg bort fra hverandre, gjør spindelstenger det også, og forlenger cellen.
På slutten av denne fasen er kromosomgruppene lokalisert i motsatte ender av den mitotiske spindelen, så hver ende av cellen er med et komplett og tilsvarende sett med kromosomer.
Telofase
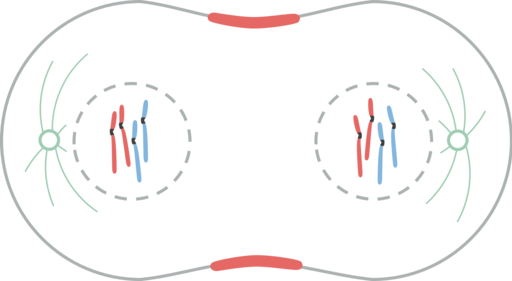 Telofase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)]
Telofase. Leomonaci98 [CC By-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] Telophase er den siste fasen av atomavdelingen. Cinetocoro -mikrotubuli går i oppløsning mens polare mikrotubuli blir ytterligere forlenget.
Atommembranen begynner å danne seg rundt hvert kromosomspill, ved å bruke kjernefysiske innpakning av overordnede celle, som var som cytoplasme -vesikler.
På dette stadiet frarådes kromosomene som er i cellepolene fullstendig på grunn av defosforylering av histonmolekyler (H1). Dannelsen av elementene i kjernemembranen er rettet av flere mekanismer.
Under anafasen begynte mange av de fosforylerte proteiner å bli defosforilert i profasen. Dette tillater begynnelsen på telofasen, at kjernefysiske vesikler begynner å montere på nytt, assosiert med overflaten av kromosomene.
Kan tjene deg: kondrocytter: Kjennetegn, histologi, funksjoner, dyrkingPå den annen side er kjernefysisk pore på nytt som tillater pumping av nukleære proteiner. Nukleære lamina -proteiner er defosforylert, slik at de kan være assosiert igjen, for å fullføre dannelsen av nevnte kjernemynett.
Til slutt, etter at kromosomene er fullstendig motløs, startes syntesen av RNA, og danner nukleolus igjen og dermed fullfører dannelsen av de nye grensesnittkjernene til datterceller.
Cytokinesis

Cytocinesis tas som en hendelse atskilt fra kjernefysisk inndeling, og ofte i typiske celler, følger den cytoplasmatiske inndelingsprosessen hver myitose, som starter i anafase. Flere studier har vist at i noen embryoer oppstår flere kjernefysiske inndeling før den cytoplasmatiske inndelingen.
Prosessen begynner med utseendet til en rille eller spalte som er merket i planet til den metafasiske plaketten, noe som sikrer at inndelingen oppstår mellom kromosomgruppene. Spaltet er indikert av den mitotiske spindelen spesifikt, mikrotubulene i Ásteres.
I den markerte kløften er det en serie mikrofilamenter som danner en ring rettet mot den cytoplasmatiske siden av cellemembranen, i stor grad sammensatt av aktin og myosin. Disse proteinene samhandler med hverandre og tillater sammentrekning av ringen rundt sporet.
Denne sammentrekningen genereres ved glidning av filamentene til disse proteinene, når de samhandler med hverandre, på samme måte som de gjør for eksempel i muskelvev.
Sammentrekningen av ringen blir utdypet ved å utøve en "klemmende" effekt som til slutt deler foreldrecellen, og tillater separasjon av datterceller, med det cytoplasmatiske innholdet i utvikling.
Cytocinesis i planteceller
Planteceller har en cellevegg, så deres cytoplasmatiske inndelingsprosess er forskjellig fra den som er beskrevet tidligere og begynner i telofase.
Dannelsen av en ny cellevegg begynner å sette sammen mikrotubuli i den gjenværende spindelen, og utgjør ramoplasten. Denne sylindriske strukturen dannes av to Microtubules -spill som kobles sammen i endene, og hvis positive stolper er innebygd i en elektronisk plakett i det ekvatoriale planet.
Små vesikler fra Golgi -apparatet, fullt av forløpere av celleveggen, beveger seg gjennom mikrotubuli av ramoplasten til ekvatorialregionen, og kombinerer for å danne en celleplakk. Innholdet i vesiklene er adskilt på denne platen når det vokser.
Denne platen vokser og smelter sammen med plasmamembranen langs celleomkretsen. Dette skjer på grunn av konstant omorganisering av ramplastens mikrotubuli i platenes periferi, slik at flere vesikler kan bevege seg mot dette planet og tømme innholdet.
På denne måten oppstår den cytoplasmatiske separasjonen av datterceller. Endelig innholdet i celleplaten ved siden av cellulosemikrofiber i den, gjør det mulig å fullføre dannelsen av den nye celleveggen.
Funksjoner
Mitose er en delingsmekanisme i celler, og er en del av en av fasene i cellesyklusen i eukaryoter. På en enkel måte kan vi si at hovedfunksjonen til denne prosessen er reproduksjon av en celle i to datterceller.
For encellede organismer betyr celledeling generering av nye individer, mens for flercellede organismer er denne prosessen en del av veksten og riktig funksjon av den komplette kroppen (celledelingen genererer utvikling av vev og vedlikehold av strukturer).
Mytosprosessen er aktivert i henhold til organismenes krav. Hos pattedyr, for eksempel, begynner røde blodlegemer (erytrocytter) å dele flere celler, når kroppen trenger en bedre oksygenfangst. Tilsvarende reproduserer hvite blodlegemer (leukocytter) når det er nødvendig å bekjempe en infeksjon.
Derimot mangler noen spesialiserte dyreceller, praktisk talt mitoseprosessen eller er veldig treg. Eksempel på dette er nerveceller og muskelceller).
Generelt er de celler som er en del av organismenes binde og strukturelle vev og hvis reproduksjon bare er nødvendig når en celle har en viss feil eller forverring og må erstattes.
Vekstregulering og celledeling.
Cellevekst- og celledelingssystemet er mye mer sammensatt i flercellede organismer enn i encellede. I sistnevnte er reproduksjon i utgangspunktet begrenset av tilgjengeligheten av ressurser.
I dyreceller blir divisjonen varetektsfengslet til det er et positivt signal som aktiverer denne prosessen. Denne aktiveringen kommer i form av kjemiske signaler fra naboceller. Dette gjør det mulig å forhindre ubegrenset vekst av vev, og reproduksjon av mangelfulle celler, som alvorlig kan skade organismenes liv.
Kan tjene deg: Basal membran: Karakteristikker, struktur og funksjonerEn av mekanismene som kontrollerer cellemultiplikasjon er apoptose, der en celle dør (på grunn av produksjon av visse proteiner som aktiverer selvdestruksjon) hvis den har betydelig skade eller er infisert av et virus.
Det er også regulering av celleutvikling gjennom hemming av vekstfaktorer (for eksempel protein). Dermed forblir cellene i grensesnittet, uten å gå videre til M -fasen av cellesyklusen.
Organismer som utfører det
Mitoseprosessen utføres i de aller fleste eukaryote celler, fra encellede organismer som gjær, som bruker den som en aseksuell reproduksjonsprosess, til komplekse flercellede organismer som planter og dyr og dyr.
Selv om cellesyklusen generelt er den samme for alle eukaryote celler, er det bemerkelsesverdige forskjeller mellom encellede og flercellede organismer. I førstnevnte er veksten og delingen av celler foretrukket av naturlig seleksjon. I flercellede organismer er spredning begrenset av strenge kontrollmekanismer.
I encellede organismer skjer reproduksjon akselerert, siden cellesyklusen fungerer konstant og datterceller raskt tar fatt på myitose for å fortsette med den syklusen. Mens cellene i flercellede organismer tar betydelig mer tid å vokse og dele seg.
Det er også noen forskjeller mellom de mitotiske prosessene til plante- og dyreceller, som i noen av fasene i denne prosessen, men i prinsippet fungerer imidlertid på samme måte i disse organismer.
Cellulær inndeling i prokaryote celler
Prokaryotisk celle Generelt vokser prokaryote celler og er delt med en raskere hastighet enn eukaryote celler.
Organismer med prokaryote celler (vanligvis encellede eller i noen flercellede tilfeller) mangler en kjernefysisk membran som isolerer det genetiske materialet inne i en kjerne, så det er spredt i cellen, i et område som kalles nukleoid. Disse cellene har et viktig sirkulært kromosom.
Celledeling i disse organismer er da mye mer direkte enn i eukaryote celler, som mangler den beskrevne mekanismen (mitose). I dem utføres reproduksjon av en prosess som kalles binær fisjon, der DNA -replikasjon begynner i et spesifikt sirkulært kromosomsted (Replication Origin eller ORIC).
Det dannes to opprinnelser som vandrer til motsatte sider av cellen når replikasjon oppstår, og cellen strekker seg til den når dobbelt så stor størrelse. På slutten av replikasjonen vokser cellemembranen inn i cytoplasma, og deler avkomcellen i to døtre med samme genetiske materiale.
Evolusjon av mitose
Utviklingen av eukaryote celler førte til økningen i kompleksiteten i genomet. Dette antydet utviklingen av mer forseggjorte divisjonsmekanismer.
Hva som gikk foran mitosen?
Det er hypoteser som foreslår at bakteriell inndeling er forgjengermekanismen for mitose. En viss sammenheng mellom proteinene assosiert med binær fisjon er funnet (som kan være de som forankrer kromosomene til spesifikke steder i plasmamembranen til døtrene) med tubulinet og aktinet til de eukaryote celler.
Noen studier indikerer visse særegenheter i delingen av moderne encellede protister. I dem forblir kjernemembranen intakt under mitose. Replikerte kromosomer forblir forankret til visse steder i denne membranen, og skiller seg når kjernen begynner å strekke seg under celledelingen.
Dette viser en viss tilfeldighet med den binære fisjonsprosessen, der replikerte kromosomer er fikset til visse steder i cellemembranen. Hypotesen foreslår da at protistene som presenterer denne kvaliteten under celledelingen, kunne ha opprettholdt dette kjennetegnet for en forfedres cellulær celle.
For tiden, forklaringer på hvorfor eukaryote celler i flercellede organismer ennå ikke er utviklet, er det nødvendig at kjernemembranen går i oppløsning under celledelingsprosessen.
Referanser
- Albarracín, a., & Telulón, til. TIL. (1993). Cellteori i det nittende århundre. Akal -utgaver.
- Alberts, f., Johnson, a., Lewis, J., Raff, m., ROBERTH, K., & Walter, P. (2008). Biologi av cellemolekylæren. Garland Science, Taylor og Francis Group.
- Campbell, n., & Reece, J. (2005). Biologi 7th Utgave, ap.
- Griffiths, a. J., Lewontin, r. C., Miller, J. H., & Suzuki, D. T. (1992). Introduksjon til genetisk analyse. McGraw-Hill Inter-American.
- Karp, g. (2009). Celle- og molekylærbiologi: konsepter og eksperimenter. John Wiley & Sons.
- Lodish, h., Darnell, J. OG., Berk, a., Kaiser, ca. TIL., Krieger, m., Scott, m. P., & Matsudaira, P. (2008). Mollekylær cellebiologi. Macmillan.
- Segura-Valdez, m. D. L., Cruz-Gómez, s. D. J., López-Cruz, r., Zavala, g., & Jiménez-García, l. F. (2008). Visualisering av mitose med atomkraftmikroskop. Tips. Spesialisert magasin i Chemical-Biological Sciences, 11 (2), 87-90.