

Nukleære poregenskaper, funksjoner, komponenter
- 4806
- 1131
- Jonathan Carlsen
Han atompore (Fra det greske, porer = trinn eller transitt) er "døren" til kjernen som tillater det meste av transporten mellom nukleoplasma og cytoplasma. Atomporen sammenføyer de indre og ytre membranene i kjernen for å danne kanaler, som tjener til transport av proteiner og RNA.
Ordet poro gjenspeiler ikke kompleksiteten i den aktuelle strukturen. På grunn av dette er det å foretrekke å referere til kjernefysisk porekompleks (CPN), i stedet for kjernefysisk pore. CPN kan oppleve endringer i strukturen under transport eller tilstander i cellesyklusen.
Nylig har det blitt oppdaget at nukleoporiner, proteiner som utgjør CPN, har en viktig rolle i å regulere genuttrykk. Når mutasjoner oppstår som påvirker funksjonen til nukleoporiner, produseres patologier hos mennesker, for eksempel autoimmun, kardiomyopatier, virusinfeksjoner og kreft.
[TOC]
Kjennetegn
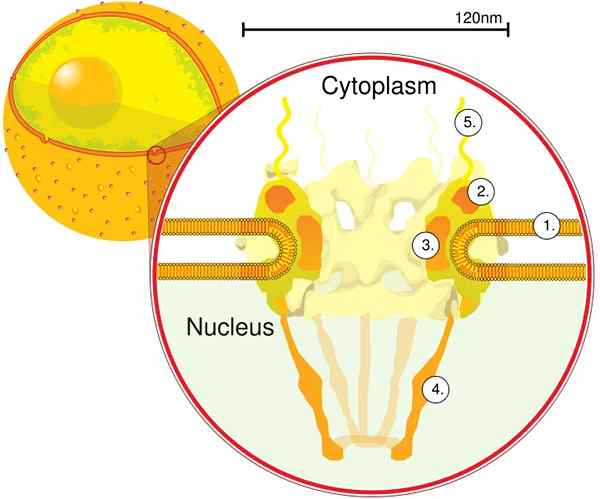
Gjennom elektronomografi ble det bestemt at CPN har en tykkelse på 50 nm, en ekstern diameter mellom 80 og 120 nm, og en indre diameter på 40 nm. Store molekyler som den store underenheten til ribosomer (PM 1590 kDa) kan eksporteres utenfor kjernen gjennom CPN. Det anslås at det er mellom 2000 og 4000 CPN per kjerne.
Molekylvekten til en individuell CPN er omtrent mellom 120 og 125 MDA (1 MDA = 106 Da) i virveldyr. I kontrast er CPN mindre i gjær, der den har omtrent 60 MDA. Til tross for den enorme størrelsen på CPN, er nukleoporiner sterkt bevart i alle eukaryoter.
Translokalisering gjennom CPN er en rask prosess, hvis hastighet er 1000 translokasjoner/sekund. Imidlertid bestemmer CPN ikke retningen på transportstrømmen i seg selv.
Dette avhenger av rangtp -gradienten, som er større i kjernen enn i cytosol. Denne gradienten opprettholdes av en guaninvekslerfaktor som kjøres.
Under cellesyklusen opplever CPNS montering og uenighetssykluser. Monteringen skjer ved grensesnittet og umiddelbart etter mitose.
Funksjoner
Ribonukleinsyre (lite nukleært RNA, messenger RNA, overføring RNA), proteiner og ribonukleoproteiner (RNP) må transporteres aktivt gjennom CPN. Dette betyr at energien til ATP- og GTP -hydrolyse er nødvendig. Hvert molekyl transporteres på en bestemt måte.
Generelt sett er RNA -molekyler fullpakket med proteindannende RNP -komplekser, og eksporteres på denne måten. I kontrast må proteiner som blir aktivt transportert til kjernen ha et sekvensskilt av plassering i kjernen (SLN), besitter av aminosyreavfall med positiv belastning (for eksempel KKKRK).
Kan tjene deg: mesosomProteiner som eksporteres til kjernen må ha et eksportsignal til kjernen (NES) rik på leucinaminosyre.
I tillegg til å tilrettelegge for transport mellom kjernen og cytoplasma, er CPN -er involvert i kromatinorganisasjon, regulering av genuttrykk og DNA -reparasjon. Nukleoporiner (NUPs) fremmer transkripsjonsaktivering eller undertrykkelse, uavhengig av celleproliferasjonstilstand.
I gjær finnes nutene i CNP for kjernefysisk innpakning. I metazoariene er de inne. De utfører de samme funksjonene i alle eukaryoter.
Import av stoffer
Gjennom CPN er det passiv formidling av små molekyler i både retninger og aktiv transport, proteinimport, eksport av RNA og ribonukleoproteiner (RNPs), og de toveismolekylene Shuttle. Sistnevnte inkluderer RNA, RNP og proteiner involvert i signalering, biogenese og erstatning.
Import av proteiner i kjernen skjer i to trinn: 1) proteinbinding til den cytoplasmatiske siden av CPN; 2) ATP -avhengig translokalisering gjennom CPN. Denne prosessen trenger ATP -hydrolyse og utveksling av GTP/BNP mellom kjernen og cytoplasmaet.
I følge en transportmodell beveger det mottakende proteinkomplekset seg langs kanalen gjennom unionen, dissosiasjon og union igjen til de gjentatte FG-sekvensene av nukleoporiner. På denne måten beveger komplekset seg fra en nukleoporin til en annen inne i CPN.
Eksport av stoffer
Ligner import. Ran GTPase pålegger transport retning gjennom CNP. RAN er en molekylær bryter med to konformasjonstilstander, avhengig av om den er koblet til BNP eller GTP.
To spesifikke regulatoriske proteiner utløser konverteringen mellom de to tilstandene: 1) cytosolisk GTPase-aktivering av protein (GAP), som produserer GTP-hydrolyse og konverterer dermed Ran-GTP til Ran-BNP; og 2) Nuclear Guanina Exchange Factor (GEF), som fremmer utveksling av BNP med GTP og konverterer Ran-BNP til Ran-GTP.
Cytosol inneholder hovedsakelig Ran-BNP. Kjernen inneholder hovedsakelig ran-gtp. Denne gradienten av de to konformasjonsformene for RAN -direktører transport i den aktuelle adressen.
Import av mottakeren, sammen med stillingen, tilrettelegges av unionen til repetisjonene-FG. Hvis du når kjernesiden av CNP, blir Ran-GTP med på mottakeren for å frigjøre din posisjon. Dermed skaper Ran-GTP retningen på importprosessen.
Nukleær eksport er lik. Ran-GTP i kjernen fremmer imidlertid foreningen av posisjonen til eksportmottakeren. Når eksportmottakeren beveger seg gjennom porene mot cytosolen, møter den Ran-Gap, som induserer GTP-hydrolyse til BNP. Til slutt frigjøres mottakeren fra sin stilling og løp-BNP i cytosol.
Kan tjene deg: polymorfonukleære leukocytterRNA -transport
Eksporten av noen slags RNA ligner eksport av proteiner. For eksempel bruker Arnt og Rnans (Small Nuclear) RangTP -gradienten og blir transportert gjennom CPN ved hjelp av midler. Eksporten av modne ribosomer er også avhengig av rangtp -gradienten.
MRNA blir eksportert på en veldig annen måte fra proteinene og andre RNA -er. For sin eksport danner mRNA et RNP Messenger (RNPM) -kompleks, der et RNM -molekyl er omgitt av hundrevis av proteinmolekyler. Disse proteinene har behandlingen, Kapping, Empalme og polyadentilly av MNA.
Cellen må kunne skille mellom RNPM med moden RNM og RNPM med umoden mRNA. MRNA, som danner RPNM -komplekset, kan ta i bruk topologier som må ombygges for transport. Før RNPM kommer inn i CPN, oppstår et kontrolltrinn utført av Tramp og eksosomproteinkomplekser.
Når moden RNPM er samlet, transporteres RPNM gjennom kanalen ved hjelp av en transportmottaker (NXF1-NXT1). Denne mottakeren trenger ATP -hydrolyse (ikke en RANGTP -gradient) for å etablere retningsbygging av RNPM, som vil nå cytoplasmaet.
Det nukleære porekomplekset og kontrollen av genuttrykk
Noen studier indikerer at CPN -komponenter kan påvirke reguleringen av genetisk uttrykk gjennom kontroll av kromatinstruktur og dens tilgjengelighet til transkripsjonsfaktorer.
I eukaryoter av nyere evolusjon er heterokromatin fortrinnsvis lokalisert på periferien av kjernen. Dette territoriet blir avbrutt av eukromatinkanaler, som opprettholdes av CPN -kjernefysisk kurv. Nuclear Basket Association med eukromatin er relatert til gentranskripsjon.
For eksempel innebærer aktivering av transkripsjon på CPN -nivå interaksjonen mellom komponentene i kjernekurven med proteiner som histonacetyltransferasesaga og RNA -eksportfaktorer.
Dermed er kjernefysisk kurv en plattform for mange vedlikeholdsgener (Renhold) sterkt transkribert og gener sterkt indusert av endringer i miljøforholdene.
Nuclear Pore and Virology Complex
Viral eukaryot celleinfeksjon avhenger av CPN. I begge tilfeller av virusinfeksjon avhenger suksessen av DNA, RNA eller RPN som passerer gjennom CPN for å oppnå det endelige målet, som er replikasjonen av viruset.
Kan tjene deg: Cytoplasmatiske inneslutninger: Hva er, egenskaper, funksjonerAPE -virus 40 (SV40) har vært en av de mest studerte modellene for å undersøke funksjonen til CPN i translokalisering i kjernen. Dette er fordi SV40 har et lite genom (5.000 baser).
Det er vist at transport av virus -DNA er forenklet proteinproteiner i viruset, som beskytter viruset til kjernen når.
Komponenter
CPN er innebygd inne i kjernefysisk konvolutt og består av mellom ca. 500 og 1000 nupts. Disse proteinene er organisert i strukturelle subkomplekser eller moduler, som samhandler med hverandre.
Den første modulen er en sentral komponent, eller ring, inne i porene i form av en sandklokke, som er begrenset av ytterligere 120 nm diameter ring på begge sider, intranukleær og cytoplasmatisk. Den andre modulen er kjernen og cytoplasma ringer (120 nm i diameter hver) som ligger rundt sandwate -formet komponent.
Den tredje modulen er åtte filamenter som er projisert fra 120 nm ringen i nukleoplasmaet og danner en kurvformet struktur. Den fjerde modulen består av filamentene som er projisert til siden av cytoplasmen.
Det Y -formede komplekset, bestående av seks NUP -er og SEH 1 og Sec 13 -proteiner, er det største og best karakteriserte komplekset til CNP. Dette komplekset er den essensielle enheten som er en del av CPN -stillaset.
Til tross for den lave likheten mellom NUPS -sekvensene, er CPN -stillaset sterkt bevart i alle eukaryoter.
Referanser
- Beck, m., Vondt, e. 2016. Nuclear Pore Complex: Forstå dens funksjon gjennom strukturell innsikt. Nature Reviews, Molecular Cell Biology, DOI: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.W. 2015. Nukleære poreproteiner og kontroll av genomfunksjoner. Gener og utvikling, 29, 337-349.
- Kabachinski, g., Schwartz, t.ELLER. 2015. Det kjernefysiske porekomplekset - struktur og funksjon på et øyeblikk. Journal of Cell Science, 128, 423-429.
- Knocknhauer, k.OG., Schwartz, t.ELLER. 2016. Det kjernefysiske porekomplekset som fleksibel og dynamisk port. Cell, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, r.W. 2014. Det multifunksjonelle kjernefysiske av kompleks: en plattform for å kontrollere genuttrykk. Nåværende mening om cellebiologi, doi: 10.1016/j.CEB.2014.02.001.
- Stawicki, s.P., Steffen, J. 2017. Republikasjon: Atomporekomplekset - en omfattende gjennomgang av struktur og funksjon. International Journal of Academic Medicine, 3, 51-59.
- Tran, e.J., Wye, s.R. 2006. Dynamisk kjernefysisk av komplekser: Livet på kanten. Cell, 125, 1041-1053.