

Proteiner g struktur, typer og funksjoner
- 2882
- 53
- Prof. Theodor Gran
De G enten Guaninnukleotidbindende proteiner, De er proteiner assosiert med plasmamembranen som tilhører en familie av "koblinger" av signaler som har viktige funksjoner i mange signaltransduksjonsprosesser i eukaryote organismer.
I litteratur beskrives G -proteiner som binære molekylære brytere, siden deres biologiske aktivitet bestemmes av endringene i deres struktur gitt av nukleotidartene som de er i stand til å sammenføye: guanosin -nukleotider (difosfat (BNP) og typosfat (GTP)).
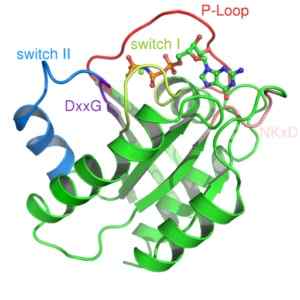
Struktur av Ras -proteinet, et monomer G -protein (kilde: Mark 'Absturz' [Public Domain] via Wikimedia Commons) De aktiveres vanligvis av reseptorer av en familie av protein kjent som GPCR -koblede reseptorer (GPCR, engelsk "G-protein-koblede mottakere "), som mottar et eksternt innledende signal og gjør det konformasjonsendringer som utløser aktivering, som deretter oversettes til aktivering av et annet effektorprotein.
Noen forfattere vurderer at kodingsgenene for denne proteinfamilien utviklet seg i duplisering og divergens av et forfedres vanlig gen, hvis produkt ble foredlet og spesialiserer mer og mer.
Blant det store utvalget av cellulære funksjoner som disse proteinene har, er translokasjonen av makromolekyler under proteinsyntese, transduksjon av hormonelle signaler og sensoriske stimuli, samt regulering av spredning og celledifferensiering.
To klasser er beskrevet fra denne typen protein: små G -proteiner og heterotrofimeriske G -proteiner. Den første tre -dimensjonale strukturen av et G -protein ble oppnådd mer enn et tiår fra et lite G -protein kjent som RAS.
[TOC]
Struktur
Strukturelt sett blir to typer G -proteiner gjenkjent: små G -proteiner og heterotrimiske G -proteiner, mye mer komplekse.
Små G-proteiner dannes av et enkelt polypeptid på rundt 200 aminoaceous avfall og omtrent 20-40 kDa, og i dets struktur er det et bevart katalytisk domene (domenet G) sammensatt av fem α-propeller, seks β-pleter ark og fem polypeptid løkker.
Heterotrofimeriske G -proteiner er derimot omfattende membranproteiner som er sammensatt av tre polypeptidkjeder, kjent som α, β og y underenheter.
-A -underenheten veier mellom 40 og 52 kDa, har et guaninnukleotidbindingsregion og har GTPASA -aktivitet til hydrolyserkoblinger mellom GTP -fosfatgrupper.
Α -underenhetene til forskjellige G -proteiner deler noen strukturelle domener som GTP -union og hydrolyse, men er veldig forskjellige i reseptorbindingsstedene og effektorproteiner.
Kan tjene deg: mitotisk spindel-Β -underenheten har en litt lavere molekylvekt (mellom 35 og 36 kDa).
-Y -underenheten er derimot mye mindre og har en omtrentlig molekylvekt på 8 kDa.
Alle heterotrofimeriske g. Disse to domenene er så sterkt assosiert at de er tenkt som en enkelt funksjonell enhet.
Folkens
Som nevnt tidligere er det to typer G -proteiner: små og heterotriméricas.
Små G -proteiner har funksjoner i cellevekst, proteinsekresjon og interaksjon av intracellulære vesikler. For deres del er heterotrofimeriske G -proteiner assosiert med signaloverføring fra overflatreseptorer, og fungerer også som brytere som veksler mellom to tilstander avhengig av det tilhørende nukleotid.
Små G -proteiner
Disse proteinene kalles også små GTPASA -er, små GTP -kryssproteiner eller Ras -protein superfamilie og danner en uavhengig superfamilie i den store klassen av hydrolase GTP med regulatoriske funksjoner.
Disse proteinene er veldig forskjellige og kontrollerer flere celleprosesser. De er preget av et bevart domene av binding til GTP, domenet "G". Foreningen av dette nukleotidfosfatet forårsaker små konformasjonsendringer i dets katalytiske domene i GD -proteiner.
Aktiviteten er intimt relatert til GTPASAS som aktiverer proteiner (gap, engelsk "GTPase aktivering av protein ") og med guanina nukleotidutvekslingsfaktorer (GEF, engelsk "Guaninnukleotidutvekslingsfaktor ”).
I eukaryoter er det beskrevet fem klasser eller familier med små G -proteiner:
-Ras
-Rho
-Rab
-SAR1/ARF
-Løp
RAS- og Rho -proteiner kontrollerer genetisk ekspresjon og Rho -proteiner modulerer også omorganiseringen av cytoskjelettet. Rab og SAR1/ARF -gruppeproteiner har innflytelse på vesikulær transport og proteiner regulerer kjernetransport og cellesyklus.
Heterotrofimeriske G -proteiner
Denne typen protein fortjener også en assosiasjon med to andre proteinfaktorer, slik at signalveien fra utsiden til det cellulære interiøret består av tre elementer i følgende rekkefølge:
- De koblede reseptorer Et proteiner g
- De G
- De Proteiner eller Effektive kanaler
Det er et stort mangfold av heterotrofimerisk G -protein, og dette er relatert til det store mangfoldet av α -underenheter som eksisterer i naturen, der bare 20% av aminosyresekvensen er bevart.
Vanligvis identifiseres heterotrofimeriske G -proteiner takket være mangfoldet av α -underenhet, hovedsakelig basert på deres funksjonelle og sekvenslikheter.
A -underenhetene utgjør fire familier (GS -familien, GI/O -familien, GQ -familien og G12 -familien). Hver familie er sammensatt av en annen "isotype" som sammen legger til mer enn 15 forskjellige måter å α -underenheter.
Familie gs
Denne familien inneholder representanter som også deltar i den positive reguleringen av syklase -adenylatproteiner og uttrykkes i de fleste celletyper. Det er sammensatt av to medlemmer: GS og golf.
Abonnementet "S " Det refererer til stimulering (fra engelsk "Stimulering ") og abonnementet "OLF " Det refererer til "lukt" (fra engelsk "Olfaction"). Golfproteiner uttrykkes spesielt i sensoriske nevroner som er ansvarlige for lukt.
Familie gI/O
Dette er den største og mangfoldige familien. De kommer til uttrykk i mange celletyper og formidler den mottakende avhengig hemming av flere typer adenylciclasses (abonnementet "Yo " refererer til hemming).
Proteiner med gruppe α -underenheter kommer til uttrykk spesielt i celler i sentralnervesystemet og har to varianter: A og B.
Familie gq
Proteiner med denne familien av α -underenhet er ansvarlige for regulering av fosfolipase C. Denne familien består av fire medlemmer hvis α -underenheter er uttrykt av forskjellige gener. De er rikelig i leverceller, i nyreceller og lunger.
Familie g12
Denne familien kommer til uttrykk allestedsnærværende i organismer, og den er ikke kjent med sikkerhet hva de cellulære prosessene reguleres gjennom proteiner med disse underenhetene er.
Β- og γ -underenheter
Selv om mangfoldet av alfa -strukturer er avgjørende for identifisering av heterotimeriske proteiner, er det også mye mangfold med hensyn til de to andre underenhetene: beta og gamma.
Funksjoner
G -proteiner deltar i "kanaliseringen" av signalene fra reseptorene i plasmamembranen til effektorkanalene eller enzymer.
Det vanligste eksemplet på funksjonen til denne typen protein er i reguleringen av enzymet adenylat cyclasa, enzym som er ansvarlig for syntesen av adenosin 3 ', 5'-monofosfat eller ganske enkelt syklisk AMP, et molekyl som har viktig funksjon som andre messenger I mange kjente celleprosesser:
Kan tjene deg: caliciform celler-Selektiv proteinfosforylering med spesifikke funksjoner
-Genetisk transkripsjon
-Omorganisering av cytoskjelettet
-Sekresjon
-Membran -depolarisering
De deltar også indirekte i reguleringen av inositolene som signaliserer foss (fosfatidylinitol og deres fosforylerte derivater), som er ansvarlige for kontroll av kalsiumavhengige prosesser som kjemiotaxis og sekresjonen av oppløselige faktorer.
Mange ioniske kanaler og transportproteiner styres direkte av proteinproteiner g. Tilsvarende er disse proteinene involvert i mange sensoriske prosesser som syn, lukt, blant andre.
Hvordan fungerer de?
Modusen for interaksjon av et G -protein med effektorproteiner er typisk for hver klasse eller familie av protein.
For G-grebranale reseptor G-proteiner (heterotrofimeriske G-proteiner), forening av et guaninkjerne som BNP eller guanosín difosfat for å underenhet α forårsaker assosiasjonen til de tre underenhetene, og danner et kompleks kjent som Gαβy eller G-GDP, som er koblet til et kompleks kjent som Gαβy eller G-GDP, som er koblet til en kompleks kjent som Gαβy eller G-GDP, som er koblet til en kompleks kjent som GαβUnit a til membranen.
Hvis BNP-molekylet deretter utveksles med et GTP-molekyl, dissosierer α-underenheten festet til GTP-dissosiatene fra β- og y-underenhetene, og danner et eget kompleks kjent som Gα-GTP, som er i stand til å endre aktiviteten til enzymer eller Diana-transportør, som er i stand til å endre aktiviteten til enzymer eller Diana-transportør, som er i stand til å endre aktiviteten til den proteiner.
Den hydrolytiske aktiviteten til denne underenheten lar deg fullføre aktiveringen, og utveksle GTP for et nytt BNP, og flytter til den inaktive konformasjonen.
I fravær av de eksiterte reseptorene som er assosiert med BNPs BNP -utvekslingsprosess, er det veldig tregt, noe som betyr at heterotrimiske GD -proteiner bare utveksler BNP med GTP med en fysiologisk viktig hastighet når de blir knyttet til deres begeistrede reseptorer.
Referanser
- Gilman, g. (1987). G proteiner: svinger av mottakergenenede signaler. Årlige anmeldelser i biokjemi, 56, 615-649.
- Milligan, g., & Kostenis, og. (2006). Heterotrimiske G-proteiner: En kort historie. British Journal of Pharmacology, 147, 546-555.
- Offermanns, s. (2003). G-proteiner som svinger i transmembran signalering. Fremgang i biofysikk og molekylærbiologi, 83, 101-130.
- Simon, m., Strathmann, m. P., & Gautam, n. (1991). Mangfold av G -proteiner i signaloverføring. Vitenskap, 252, 802-808.
- Syrovatkina, v., Munter, k. ENTEN., Dey, r., & Huang, x. (2016). Regulering, signalering og fysiologiske funksjoner av G-proteiner. Journal of Molecular Biology, 428(19), 3850-3868.
- « Crotalus triseriatus -egenskaper, habitat, reproduksjon
- Tordenegenskaper, habitat, taksonomi, egenskaper »