

Riboseegenskaper, struktur og funksjoner
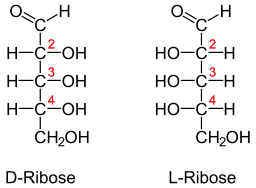
- 2480
- 535
- Daniel Skuterud
De Ribosa Det er et fem -karbon sukker som er til stede i ribonukleosider, ribonukleotider og dets derivater. Du kan finne andre navn som β-d-librounosa, d-libose og l-libose.
Nukleotider er de konstituerende "blokker" av ribonukleinsyreskjelettet (RNA). Hvert nukleotid er sammensatt av en base som kan være adenin, guanin, cytosin eller uracil, en fosfatgruppe og et sukker, ribosen.
Fisher-projeksjon for D- og L-Libose (Kilde: Neurotokeker [Public Domain] via Wikimedia Commons) Denne typen sukker er spesielt rikelig i muskelvev, der det er assosiert med ribonukleotider, spesielt med adenosin eller ATP -tryfosfat, noe som er essensielt for muskelfunksjon.
D-ribose ble oppdaget i 1891 av Emil Fischer, og siden har det blitt lagt stor vekt på dens fysisk-kjemiske egenskaper og dens rolle i cellemetabolismen, det vil si som en del av skjelettet til ribonukleinsyre, ATP og flere av flere koenzymer.
I begynnelsen ble dette bare oppnådd fra hydrolyse av RNA fra gjær, inntil det på 1950-tallet klarte å syntetisere fra D-glukosen i mer eller mindre tilgjengelige mengder, noe som tillot industrialisering av produksjonen.
[TOC]
Kjennetegn
Ribosen er en ofte ekstrahert aldopentosa som en ren kjemisk forbindelse i form av d-libose. Det er et vannløselig organisk stoff, hvitt og krystallinsk. Å være et karbohydrat, har ribose polare og hydrofile egenskaper.
Ribosen er i samsvar med den vanlige regelen for karbohydrater: den har samme antall karbon- og oksygenatomer, og det dobbelte av denne mengden i hydrogenatomer.
Gjennom karbonatomer i posisjoner 3 eller 5, kan dette sukkeret bli sammen med en fosfatgruppe, og hvis det blir sammen med en av de nitrogene basene til RNA, dannes et nukleotid et nukleotid.
Den vanligste måten å finne ribose i naturen er som d-libose og 2-deexy-d-libose, dette er komponenter av nukleotider og nukleinsyrer. D-libose er en del av ribonukleinsyre (RNA) og 2-Disaxi-D-dibosen av deoksyribonukleinsyre (DNA).
Kan tjene deg: Protein K: Kjennetegn, enzymatisk aktivitet, applikasjoner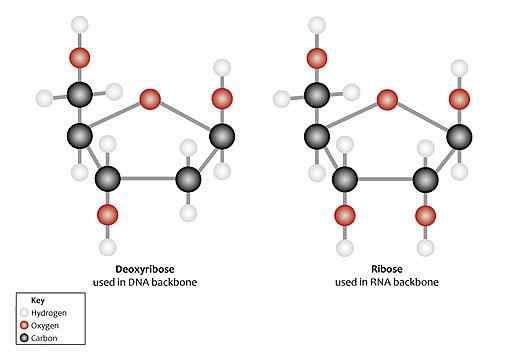 Strukturelle forskjeller mellom ribose og deoksyribose (Kilde: Genomics Education Program [CC av 2.0 (https: // creativecommons.Org/lisenser/av/2.0)] via Wikimedia Commons)
Strukturelle forskjeller mellom ribose og deoksyribose (Kilde: Genomics Education Program [CC av 2.0 (https: // creativecommons.Org/lisenser/av/2.0)] via Wikimedia Commons) I nukleotider finnes begge typer pentose i ß-fluanøs form (lukket femkantet ring).
I løsning er fri ribose i likevekt mellom aldehyd (åpenkjede) og den ß-furanøse sykliske form. Imidlertid inneholder RNA bare β-D-dribofuranosa syklisk form. Den biologisk aktive formen er vanligvis D-libosen.
Struktur
La Ribosa er et sukker avledet fra glukose som tilhører Aldopentosa -gruppen. Den molekylære formelen er C5H10O5 og har en molekylvekt på 150.13 g/mol. Siden det er et monosakkaridsukker, skiller hydrolysen molekylet i sine funksjonelle grupper.
Den har, som formelen indikerer, fem karbonatomer som kan finnes syklisk som en del av fem eller seks medlemsringer. Dette sukkeret har en aldehydgruppe i karbonatom 1 og en hydroksylgruppe (-OH) i karbonatomer fra posisjon 2 til posisjon 5 av pontosa-ringen.
Ribosemolekylet kan være representert i projeksjonen av Fisher på to måter: d-libose eller l-libose, og er formen l stereoisomeren og enantiomer i formen d og omvendt.
Klassifiseringen av D- eller L -formen avhenger av orienteringen til hydroksylgruppene i det første karbonatom etter aldehydgruppen. Hvis denne gruppen er orientert mot høyre side, tilsvarer molekylet på vegne av Fisher D-Riby.
Haworth -projeksjonen av ribosen kan representeres i to ekstra strukturer avhengig av orienteringen til hydroksylgruppen i karbonatomet som er anomer. I ß -posisjonen er hydroksylen orientert mot den øvre delen av molekylet, mens α -posisjonen guider hydroksyl mot bunnen.
Det kan tjene deg: Flora og Fauna of Santa Fe: Representative arter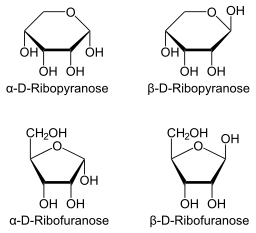 Haworth -projeksjon for ribopyranosa og ribofuranosa (kilde: Neurotokeker [Public Domain] via Wikimedia Commons)
Haworth -projeksjon for ribopyranosa og ribofuranosa (kilde: Neurotokeker [Public Domain] via Wikimedia Commons) I henhold til Haworth-projeksjonen kan det tas fire mulige måter: β-D-libose, α-D-libose, β-L-libose eller α-l-libose.
Når fosfatgrupper blir koblet til ribosen, kalles de vanligvis α, β og ƴ. Hydrolyse av nukleosider tryfosfat gir kjemisk energi for å øke et bredt utvalg av celleaksjoner.
Funksjoner
Det er foreslått at ribosefosfatproduktet av nedbrytningen av ribonukleotidene, er en av hovedforløperne for furano og tiofenolene, som er ansvarlige for den karakteristiske lukten av kjøttet.
I celler
Den kjemiske plastisiteten til ribosen får molekylet til å være involvert i de aller fleste biokjemiske prosesser inne i celleinteriør, noen som oversettelsen av DNA, syntese av aminosyrer og nukleotider, etc.
Ribosen fungerer kontinuerlig som et kjemisk kjøretøy inne i cellen, siden nukleotidene kan presentere en, to fosfatgrupper forenet kovalent med hverandre av vannfrie bindinger. Disse er kjent som henholdsvis mono-, di- og trifosfat.
Koblingen mellom ribosen og fosfatet er av estertypen, hydrolysen av denne koblingen frigjør omtrent 14 kJ/mol under standardbetingelser, mens den for hver av de anhydrerte bindinger frigjør omtrent 30 kJ/mol.
I ribosomene kan for eksempel 2'-hydroksylgruppen av ribosen danne en hydrogenbinding med de forskjellige aminosyrene, forening som tillater proteinsyntese fra tRNA i alle kjente levende organismer.
Giften av de fleste slanger inneholder fosfodiesterase som hydrolyserer nukleotider fra 3 'enden som har en fri hydroksyl, og bryter fagforeningene mellom hydroksyl 3' av ribose eller dexyribosa.
Det kan tjene deg: Aerobia glykolyse: Hva er, reaksjoner, glykolytiske mellommennI medisin
I medisinske sammenhenger brukes det til å forbedre ytelsen og treningskapasiteten ved å øke muskelenergien. Kronisk utmattelsessyndrom blir også behandlet med dette sakkaridet, så vel som fibromyalgi og visse sykdommer i koronararterien.
I forebyggende termer brukes det for å unngå muskeltretthet, kramper, smerter og stivhet etter trening hos pasienter med arvelig lidelse av myoadenylatdeaminasemangel eller AMP -mangel på AMP deaminase.
Referanser
- Alberts, f., Johnson, a., Lewis, J., Morgan, d., Raff, m., Roberts, k., & Walter, P. (2015). Biologi av cellemolekylæren (6. utg.). New York: Garland Science.
- Angyal, s. (1969). Sammensetningen og konformasjonen av sukkerarter. Angewandte Chemie - International Edition, 8(3), 157-166.
- Foloppe, n., & Mackerell, a. D. (1998). Konformasjonsegenskaper for deoksyribose og ribosedeler av nukleinsyrer: en kvantemekanisk studie, 5647(98), 6669-6678.
- Garrett, r., & Grisham, C. (2010). Biokjemi (4. utg.). Boston, USA: Brooks/Cole. Cengage Learning.
- GUTTMAN, f. (2001). Nukleotider og nukleosider. Akademisk presse, 1360-1361.
- Mathews, c., Van Holde, K., & Ahern, K. (2000). Biokjemi (3. utg.). San Francisco, California: Pearson.
- Mottram, d. S. (1998). Smaksdannelse i kjøtt- og kjøttprodukter: En gjennomgang. Matkjemi, 62(4), 415-424.
- Nechamkin, h. (1958). Underinteressens etmologiske avledninger av kjemisk terminologi. Kjemisk terminologi, 1-12.
- Nelson, d. L., & Cox, m. M. (2009). Lehninger prinsipper for biokjemi. Omega -utgaver (5. utg.). https: // doi.org/10.1007/S13398-014-0173-7.2
- Shapiro, r. (1988). Prebiotisk ribosesyntese: En kritisk analyse. Opprinnelse av liv og evolusjon av biosfæren, 18, 71-85.
- Merck -indeksen på nettet. (2018). Hentet fra www.RSC.org/merck-index/monografi/m9598/dribar?q = Uautorisert
- Waris, s., Pischetsrianer, m., & Salemuddin, m. (2010). DNA -skade ved ribose: hemming ved høye ribosekonsentrasjoner. Indian Journal of Biochemistry & Biophysics, 47, 148-156.
- Webmd. (2018). Hentet 11. april 2019, fra www.Webmd.com/vitaminer/ai/ingrediensenmono-827/ribose
- Wulf, p., & Vandamme, og. (1997). Mikrobiell syntese av D-libose: metabolsk deregulering og gjæringsprosess. Fremskritt i anvendt mikrobiologi, 4, 167-214.
- Xu, z., Sha, og., Liu, c., Li, s., Liang, J., Zhou, J., & Xu, h. (2016). L -riboseisomerase og mannose -6 -fosfatisomerase: egenskaper og applikasjoner for l -riboseproduksjon. Applied Microbiology and Biotechnology, 1-9.