

Syntese av lipidtyper og hovedmekanismer
- 3752
- 1030
- Thomas Karlsen
De Lipidsyntese Den består av en serie enzymatiske reaksjoner ved hjelp av hvilke kortkjedehydrokarboner blir kondensert for å danne lengre kjedemolekyler som deretter kan lide forskjellige kjemiske modifikasjoner.
Lipider er en veldig variert type biomolekyler syntetisert av alle levende celler og er spesialisert i flere essensielle funksjoner for å opprettholde cellelivet.
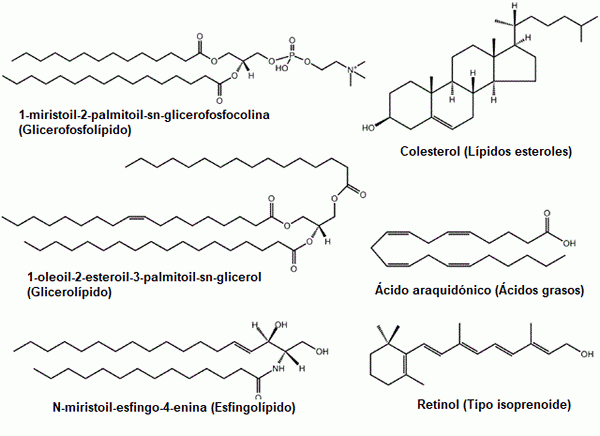 Noen eksempler på vanlige lipider: glyserofosfolipider, steroler, glyserolipider, fettsyrer, sfingolipider og prenoler (kilde: den opprinnelige opplasteren var lmaps på engelsk wikipedia. / GFDL 1.2 (http: // www.gnu.Org/lisenser/old-lisens/fdl-1.2.HTML) via Commons, tilpasset av Raquel Parada)
Noen eksempler på vanlige lipider: glyserofosfolipider, steroler, glyserolipider, fettsyrer, sfingolipider og prenoler (kilde: den opprinnelige opplasteren var lmaps på engelsk wikipedia. / GFDL 1.2 (http: // www.gnu.Org/lisenser/old-lisens/fdl-1.2.HTML) via Commons, tilpasset av Raquel Parada) Lipider er hovedkomponentene i biologiske membraner, et faktum som gjør dem grunnleggende molekyler for eksistensen av celler som isolerte enheter i omgivelsene.
Noen lipider har også spesialiserte funksjoner som pigmenter, kofaktorer, transportører, vaskemidler, hormoner, intra-ekstracellulære budbringere, kovalente ankere for membranproteiner, etc. Derfor er evnen til å syntetisere forskjellige typer lipider kritisk for overlevelse av alle levende organismer.
Denne store gruppen av forbindelser er tradisjonelt klassifisert i flere kategorier eller undergrupper: fettsyrer (mettede og umettet), glyseridkomplekslipider (lipoproteiner).
[TOC]
Typer lipider og deres viktigste syntesemekanismer
Alle reaksjonssekvenser av lipidbiosynteseruter er endogoniske og reduktive. Med andre ord, de bruker alle ATP som en energikilde og en redusert elektrontransportør, for eksempel NADPH, som en reduserende kraft.
Deretter vil hovedreaksjonene i de biosyntetiske rutene til hovedtyper av lipider bli beskrevet, det vil si av fete og eikosanoid syrer, triacylglyceroles og fosfolipider og steroler (kolesterol) (kolesterol).
- Fettsyresyntese
Fettsyrer er ekstremt viktige molekyler fra lipid synspunkt, ettersom de er en del av de mest relevante lipidene i celler. Hans syntese, i motsetning til hva mange forskere trodde under de første studiene, består ikke av den omvendte ruten til deres ß-oksidasjon.
Faktisk forekommer denne metabolske ruten i forskjellige cellekommer og krever deltakelse av en mellomledd av tre karbonatomer kjent som malonyl-CoA, noe som ikke er nødvendig ved oksidasjon.
 Malonyl-Coa. Neuroteker / Pub Domain
Malonyl-Coa. Neuroteker / Pub Domain I tillegg er det nært beslektet med sulfhydrilgruppene av proteiner kjent som acylgruppetransportører (ACP, engelsk Acylbærerproteiner).
Generelt er syntesen av fettsyrer, spesielt den for den lange kjeden, en sekvensiell prosess der fire trinn gjentas i hver "retur", og under hver retur er det en mettet syregruppe som er underlaget for den neste, som som som innebærer en annen kondens med et nytt malonyl-CoA-molekyl.
I hver sving eller reaksjonssyklus strekker fettsyrekjeden to karbonatomer, til en lengde på 16 atomer (palmitat) når, hvoretter syklusen blader.
Malonyl-CoA-formasjon
Denne mellomledningen av tre karbonatomer er irreversibelt dannet fra acetyl-CoA takket være virkningen av et acetyl-CoA karboksylaseenzym, som har en biotin-prostategruppe som er kovalent knyttet til enzymet og som deltar i denne katalysen i denne katalysen i to trinn.
I denne reaksjonen blir en karboksylgruppe avledet fra et bikarbonatmolekyl (HCO3-) overført til biotinet i en ATP-avhengig form, der biotinylgruppen oppfyller funksjonen til "midlertidig transportør" av molekylet mens overføringer til acetyl -coa, Produserer malonyl-CoA.
Kan tjene deg: Oksidasetest: Foundation, Procedure and UsesI sekvensen av syntese av fettsyrer er reduksjonsmiddelet som brukes NADPH, og de aktiverende gruppene er to TIOL-grupper (-SH) som er en del av et multienzimatisk kompleks kalt syntase-fettsyre, som er den som er viktigst i katalysesyntetisk.
Hos virveldyr er synthase -fettsyrekomplekset en del av en enkelt stor polypeptidkjede, der de 7 enzymatiske aktivitetene som er karakteristisk for syntese -ruten er representert, så vel som den hydrolytiske aktiviteten som er nødvendig for å frigjøre mellommenn på slutten av syntesen.
 Syntasa fettsyreenzymstruktur (kilde: Boehringer Ingelheim/CC By-SA (https: // CreativeCommons.Org/lisenser/by-SA/4.0) via Wikimedia Commons)
Syntasa fettsyreenzymstruktur (kilde: Boehringer Ingelheim/CC By-SA (https: // CreativeCommons.Org/lisenser/by-SA/4.0) via Wikimedia Commons) De 7 enzymatiske aktivitetene til dette komplekset er: Acilo Group Transport Protein (ACP), Acetyl-CoA-ACP-transacetilas Cetoacil-ACP-reduktase (KR), β-Hydroxyacil-ACP dehydratasa (HD) og sinne-ACP-reduktase (ER).
Før kondensasjonsreaksjonene kan oppstå for å sette sammen fettsyrekjeden, blir de to TiOL -gruppene i det enzymatiske komplekset "lastet" med acylgruppene: Først av alt blir en acetyl -coA overført til -SH -gruppen til en cystein I ß-cetoacyl-ACP-syntasen av komplekset, reaksjon katalysert av acetyl-CoA-ACP-transacetyla-enzymet (AT) (AT).
Deretter overføres en malonisk gruppe fra et malonyl -CoA -molekyl til -SH -gruppen i Acilo Group (ACP) -transportøren er en del av fettsyrekomplekssyntasen.
Sekvensen av fire reaksjoner for hver "retur" av reaksjonssyklusen er som følger:
- Kondensasjon: Acetyl- og malonylgrupper "lastet" i enzymet blir kondensert for å danne et acetoacetyl -ACP -molekyl, som er koblet til ACP -delen gjennom en gruppe -SH. I dette trinnet er det et CO2-molekyl og katalyseres av ß-Zoacyl-ACP-syntasen (acetylgruppen inntar plasseringen av "terminal metyl" av acetoacetyl-ACP-komplekset).
- Reduksjon av karbonylgruppen: Karbonylgruppen i C3-posisjonen til acetoacetyl-ACP reduseres til å danne D-ß-hydroxibutiril-ACP, reaksjon katalysert av β-cetoacyl-ACP-reduktase, som NADPH bruker AS-elektron-donor.
- Dehydrering: C2 og C3 karbonatomer i D-ß-hydroxibutiril-ACP er blottet for vannmolekyler, og danner en dobbeltbinding som ender med produksjonen av den nye forbindelsen trans-∆2-butenoil-acp. Denne prosessen er mediert av et ß-hydroksyacyl-ACP dehydratase (HD) enzym (HD).
- Dobbeltbindingsreduksjon: Dobbeltbindingen til forbindelsen dannet i dehydreringstrinnet er mettet (redusert) for å gi opphav til butiril-ACP av reaksjonen katalysert av enzymet sinne-ACP-reduktase (ER), som også bruker NADPH som et reduserende middel.
Synteseaksjonene oppstår inntil et palmitatmolekyl (16 karbonatomer) er dannet, som er hydrolysert av det enzymatiske komplekset og frigjør del av endoplasmatisk retikulum og i mitokondriene.
Det kan tjene deg: Fauna og flora i det peruanske havDe andre modifikasjonene som disse molekylene kan lide som for eksempel deaturasjonene, blir katalysert av forskjellige enzymer, som vanligvis forekommer i den glatte endoplasmatiske retikulum.
- Syntese av eikosanoider
Eikosanoider er cellelipider som har funksjoner som "kort -range" messenger molekyler, produsert av noen stoffer for å kommunisere med sine nærliggende stoffer. Disse molekylene syntetiseres fra flerumettede fettsyrer av 20 karbonatomer.
Prostaglandiner
Som respons på en hormonell stimulans angriper fosfolipaseenzymet membranfosfolipider og frigjør araquidonato fra glyserol karbon 2. Denne forbindelsen omdannes til prostaglandiner takket være et enzym av den glatte endoplasmatiske retikulum med bifunksjonell aktivitet: cyclooxygenase (Cox) eller prostaglandin H2 -syntase.
Tromboxan
Prostaglandiner kan konverteres til tromboxaner takket være tromboxan synthaseful i blodplatene (trombocytter). Disse molekylene deltar i de første trinnene for blodkoagulasjon.
- Syntese av triacylglyseroler
Fettsyrer er grunnleggende molekyler for syntese av andre mer komplekse forbindelser i celler, så som triacylglyseroler eller lipidglyserofosfolipid lipidmembran (prosesser som er avhengig av cellulære metabolske behov).
Dyr produserer triacylglyseroler og glyceofosfalipider fra to vanlige forløpere: fett-CoA-acyl og 3-fosfat L-glyserol. Fett-CoA-acyl produseres av acyl-CoA-synteser som deltar i ß-oksidasjon, mens 3-fosfat L-glyserol oppnås fra glykcolyse og ved handling av to alternativ.
Triacylglyceroles dannes ved reaksjonen mellom to molekyler Flo-CoA ACIL og et 3-fosfat diacylglycerolmolekyl; Disse overføringsreaksjonene katalyseres av spesifikke transferaser.
I denne reaksjonen er det i utgangspunktet fosfatidsyre, som defosforyleres av et enzym fosfatidsyrefosfatase for å produsere 1,2-diacylglycerol, som igjen er i stand til å akseptere en tredje molekyl med fett-CoA-acyl, produserer trielcerceroll.
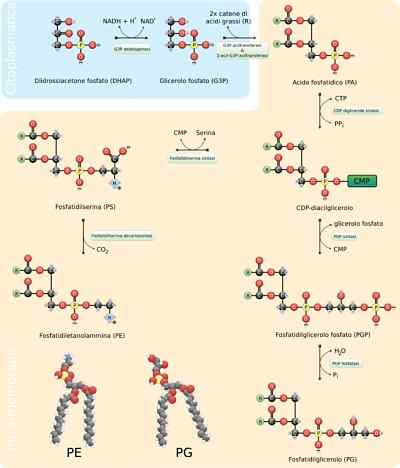
- Syntese av fosfolipider
Fosfolipider er ekstremt variable molekyler, ettersom mange forskjellige kan dannes ved kombinasjonen av fettsyrer og "hode" -grupper med glyserolskjelettet (glyceophospholipider) eller sfinosin (sfingolipider) som kjennetegner dem.
Generalforsamlingen for disse molekylene krever syntese av skjelettet til glyserol eller sfærisk, for unionen med de tilsvarende fettsyrene, enten ved sterifisering eller amidering, tilsetning av en hydrofil "hode" -gruppe gjennom en fosfodisterbinding og ,, om nødvendig , endring eller utveksling av disse siste gruppene.
I eukaryoter skjer denne prosessen i den glatte endoplasmatiske retikulumet og også i den indre mitokondrielle membranen, hvor de kan forbli på ubestemt tid eller der de kan translokeres til andre steder.
Reaksjonstrinn
De første trinnene i syntesereaksjonen av glyserofosfolipider tilsvarer de av triacylglyserolproduksjonen, som et 3-fosfat glyserolmolekyl er esterifisert til to fettsyremolekyler i karbonatomer 1 og 2, og danner fosfatidisk syre. Det er vanlig å finne fosfolipider som har mettede fettsyrer i C1 og umettet i C2 i glyserol.
Kan tjene deg: ResistinaFosfatidsyre kan også produseres ved fosforylering av et diacylglyserolmolekyl som allerede er syntetisert eller "resirkulert".
"Hodet" polare grupper av disse molekylene dannes gjennom fosfodiéster -koblinger. Det første som skal skje for at denne prosessen skal være riktig, er "aktiveringen" av en av hydroksylgruppene som deltar i prosessen med midler hydroksyl som deltar i reaksjonen.
Hvis dette molekylet binder seg til diacylglycerol, dannes CDP-diacylglycerol (den "aktiverte" formen til fosfatidsyre), men dette kan også skje på hydroksylgruppen til gruppen "hode".
Når det.
Hvis et MMP-molekyl (cytidinmonofosfat) beveger seg gjennom et nukleofilt angrep av serin eller hydroksylhydroksyl i karbonet 1 av 3-fosfatglyserol. Fosfat produserer fosfatidylglycerol.
Begge molekyler produsert på denne måten fungerer som forløpere for andre membranlipider, som ofte deler biosyntetiske ruter fra hverandre.
- Kolesterolsyntese
Kolesterol er et essensielt molekyl for dyr som kan syntetiseres av cellene, så det er ikke viktig i det daglige kostholdet. Dette molekylet med 27 karbonatomer produseres fra en forløper: acetat.
Dette komplekse molekylet er dannet fra acetyl-CoA i fire hovedtrinn:
- Kondensasjon av tre acetatenheter for å danne mevalonato, et mellomliggende molekyl med 6 karbon COA) (HMG-CoA Syntheta-enzym). Mevalonato er dannet fra HMG-CoA og takket være HMG-CoA-reduktaseenzymet.
- Mevalonato konvertering til isoprenenheter. De første 3 fosfatgruppene blir overført fra 3 ATP -molekyler til Mevalonato. En av fosfatene går tapt sammen med den tilstøtende karbonylgruppen og dannes
- Polymerisasjon eller kondensasjon av 6 isoprenenheter med 5 karbonatomer for å danne 30 -karbonatomer (et lineært molekyl) barbering (et lineært molekyl).
- Cicration of the Escualean for å danne de 4 ringene til steroidkjernen i kolesterol og påfølgende kjemiske endringer: oksidasjoner, migrasjon og eliminering av metylgrupper, etc., Hva som gir kolesterol.
Referanser
- Garrett, r. H., & Grisham, C. M. (2001). Prinsipper for biokjemi: med et menneskelig fokus. Brooks/Cole Publishing Company.
- Murray, r. K., Granner, d. K., Mayes, s. TIL., & Rodwell, V. W. (2014). Harpers illustrerte biokjemi. McGraw-Hill.
- Nelson, d. L., Lehninger, a. L., & Cox, m. M. (2008). Lehninger prinsipper for biokjemi. Macmillan.
- Jacquemyn, J., Cascaho, a., & Goodchild, r. OG. (2017). Inn og outs for endoplasmatisk retikulum - kontrollert lipidbiosyntese. Embo Reports, 18 (11), 1905-1921.
- Ohlrogge, J., & Bla gjennom, J. (nitten nitti fem). Lipidbiosyntese. Plantcellen, 7 (7), 957.
- « José López Portillo -biografi, regjering og bidrag
- Tropisk jungel i Mexico -egenskaper, beliggenhet, lettelse, flora, fauna »