

Succinato dehydrogenase struktur, funksjon, regulering, sykdommer
- 638
- 131
- Prof. Theodor Gran
SUccinato dehydrogenase (Sdh), også kjent som den komplekse II av elektrontransportkjeden, er det et mitokondrielt proteinkompleks med enzymatisk aktivitet som fungerer både i Krebs Cycle og i elektrontransportørkjeden (cellepust).
Det er et enzym som er til stede i alle aerobe celler. I eukaryoter er det et komplekst som er nært assosiert med den indre mitokondrielle membranen, mens det i prokaryotene finnes i plasmamembranen.
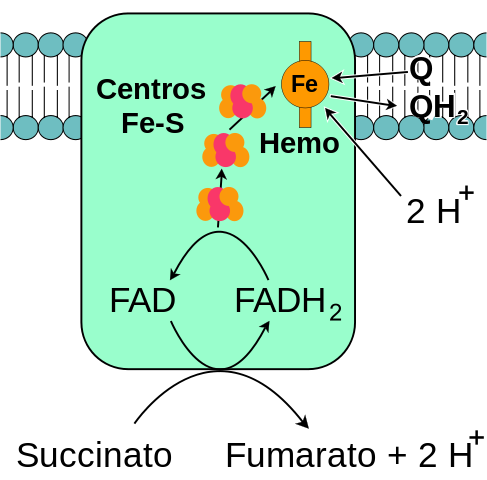 Generelt skjema for mitokondriell kompleks Succinatdehydrogenase (kilde: meg selv, basert på Fvasconcellos vektorisering. / Offentlig domene, via Wikimedia Commons)
Generelt skjema for mitokondriell kompleks Succinatdehydrogenase (kilde: meg selv, basert på Fvasconcellos vektorisering. / Offentlig domene, via Wikimedia Commons) Den komplekse succinatet dehydrogenase, oppdaget rundt 1910 og renset for første gang i 1954 av sanger og Kearney, har blitt grundig studert av flere grunner:
- Det fungerer både i Krebs -syklusen (sitronsyresyklus eller trikarboksylsyresyklus) og i elektrontransportkjeden (katalyserer oksidasjonen av succinat til fumarat)
- Aktiviteten er regulert av forskjellige aktivatorer og hemmere og
- Det er et kompleks assosiert med: jern som ikke er knyttet til en hemokruppe, labyl svovel og dyukleotider av Flavina Adenina (FAD)
Det er kodet av det nukleære genomet, og det er bevist at mutasjoner i de fire genene som kodifiserer hver av underenhetene (A, B, C og D) resulterer i forskjellige kliniske bilder, det vil si at de kan være ganske negative fra punktet av menneskers fysiske integritet.
[TOC]
Struktur
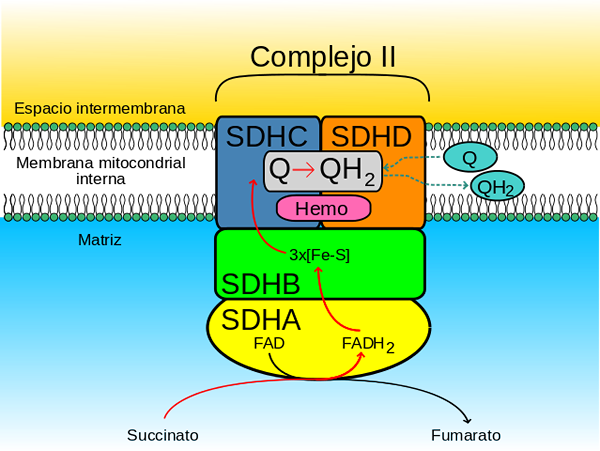
Enzymkomplekset succinatdehydrogenase dannes av fire underenheter (heterotarámero) kodet av det nukleære genomet, så det er det eneste komplekset av oksidativ fosforylering i elektrontransportørkjeden som ikke har noen underenhet kodet av den mitokondriske genom.
I tillegg er dette komplekset det eneste som ikke pumper protoner gjennom den indre mitokondrielle membranen under dens katalytiske virkning.
I henhold til studier basert på det enzymatiske komplekset av svinehjerteceller, består den komplekse succinatdehydrogenase av:
- a "hode"" Hydrofil som strekker seg fra den indre mitokondrielle membranen til mitokondriell matrise og
- a "linje"" Hydrofob som er innebygd i den indre mitokondrielle membranen og som har et lite segment som er projisert mot det oppløselige intermembranområdet til mitokondriene
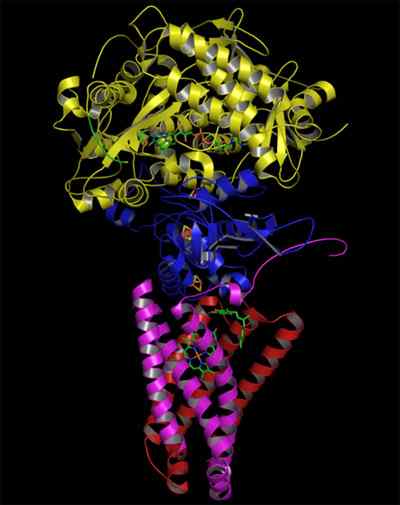
 Struktur av det komplekse succinatet dehydrogenase (kilde: Zephyris på det engelske språket Wikipedia/CC By-SA (http: // creativecommons.Org/lisenser/by-SA/3.0/) via Wikimedia Commons)
Struktur av det komplekse succinatet dehydrogenase (kilde: Zephyris på det engelske språket Wikipedia/CC By-SA (http: // creativecommons.Org/lisenser/by-SA/3.0/) via Wikimedia Commons) Hydrofil porsjonsstruktur
Det hydrofile hodet er sammensatt av SDHA (70 kDa) og SDHB (27 kDa) (SDH1 og SDH2 i gjær), og dette omfatter det katalytiske sentrum av komplekset.
SDHA- og SDHB -underenheter inneholder redoks -kofaktorer som deltar i overføring av elektroner til ubiquinon (koenzym Q10, et molekyl som transporterer elektroner mellom respirasjonskomplekser I, II og III).
SDHA-underenheten har en FAD-kofaktor (et koenzym som deltar i oksydreduksjonsreaksjoner) sammen med dens struktur, bare på krysset for succinat (hovedunderlaget til enzymet).
SDHB-underenheten har 3 jernsukkersentre (FE-S) som formidler overføringen av elektroner til ubiquinon. Et av sentrene, 2fe-2s, ligger i nærheten.
Kan tjene deg: fylogeniDet skal bemerkes at strukturelle studier indikerer at SDHB -underenhet danner grensesnittet mellom det hydrofile katalytiske domenet og domenet til "anker" til membranen (hydrofob) av komplekset.
Hydrofob porsjonsstruktur
Det som nevnt membranalt domen.
Dette domenet inneholder en hemo -del b sammen med grensesnittet mellom SDHC- og SDHD -underenhetene, der hver av de to histidinligander som holder dem sammen.
I dette enzymet er to unionssteder for Ubiquinona blitt påvist: en av stor affinitet og en av lav affinitet.
Nettstedet med høy affinitet, kjent som QP (P av proksimal) Det er ansikt til mitokondriell matrise og dannes av spesifikke aminosyrerester lokalisert i SDHB-, SDHC- og SDHD -underenheter.
Nettstedet med lav affinitet, også kalt QD (D av distalt) Det er, i den delen av den indre mitokondrielle membranen der komplekset settes inn, nærmere intermembranområdet, det vil si lenger fra organellmatrisen.
Sammen har det totale komplekset en molekylvekt nær 200 kDa, og det er bestemt at det har et forhold på 4.2-5.0 flavin nanomoler per milligram protein og 2-4 g jern for hver mol flavina.
Funksjon
Den enzymatiske succinatkompleksdehydrogenase oppfyller en viktig funksjon i mitokondriene, da ikke bare deltar i Krebs -syklusen (der den deltar i nedbrytningen av acetyl -CoA), men også en del av luftveiene, grunnleggende for energiproduksjonen ATP -formet.
Det er med andre ord et nøkkelenzym for mellomliggende metabolisme og ATP aerob produksjon.
- Det er ansvarlig for oksidasjon av succinatet til fumarat i sitronsyresyklusen
- Den mater den komplekse III av elektrontransportørkjeden med elektronene avledet fra oksidasjon av suksinatet, noe som hjelper til med å redusere oksygen og danne vann
- Elektrontransport genererer en elektrokjemisk gradient gjennom den indre mitokondrielle membranen, som favoriserer ATP -syntese
Som et alternativ kan elektroner brukes til å redusere molekyler fra et "basseng" i Ubiquinonas, og produsere de nødvendige reduksjonsmidlene for å redusere superoksydanioner som stammer fra den samme respirasjonskjeden eller som kommer fra eksogene kilder.
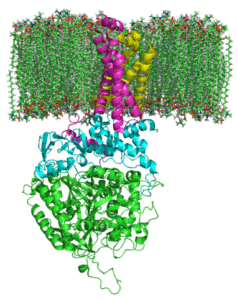
 Succinate Complex Degidrogenasa (Kilde: JohnHFST / Public Domain, via Wikimedia Commons)
Succinate Complex Degidrogenasa (Kilde: JohnHFST / Public Domain, via Wikimedia Commons) Hvordan fungerer det?
Underenheten A av komplekset (som er kovalent knyttet til FAD -koenzymet) forbinder underlag, fumarat og suksinat, så vel som dets fysiologiske regulatorer, oksalacetat (konkurrerende hemmer) og ATP.
ATP fortrenger unionen mellom oksalacetat og SDH -komplekset, og deretter blir elektronene som er "forbi" fra succinatet til SDHA -underenheten overført til gruppene av jern- og svovelatomer som er til stede i SDHB -underen ved hjelp av koenzymet FAD.
Kan tjene deg: myosin: Kjennetegn, struktur, typer og funksjonFra underenhet B når disse elektronene hemo -setene b av SDHC- og SDHD -underenhetene, hvor de blir "levert" til Quinonas -koenzymer gjennom sine unionssteder til Quinonas.
Den elektroniske strømmen fra succinatet gjennom disse transportørene og til og med den endelige akseptoren, som er oksygen, er koblet til syntesen av 1.5 ATP -molekyler for hvert elektronisk dreiemoment gjennom luftveisfosforylering.
Inzima -defekter
Det er rapportert at mutasjoner i genet som koder for underenheten A i den komplekse succinatdehydrogenase kan forårsake encefalopatier i barndommen, i mellomtiden har mutasjonene i genene som koder for underenheter B, C og D blitt assosiert med dannelsen av svulster.
Regulering
Aktiviteten til den komplekse succinatdehydrogenase kan reguleres ved post -translasjonelle modifikasjoner som fosforylering og acetylering, Selv om inhibering av aktiv sted også kan oppstå.
Acetylering av noe lysinavfall kan redusere aktiviteten til dette enzymet, og denne prosessen utføres av et enzym av acetylase kjent som SIRT3; Fosforylering har samme effekt på enzymet.
I tillegg til disse modifikasjonene, er SDH -komplekset også regulert av formidlere av Krebs -syklusen, nærmere bestemt Oksalacetat og succinat. Oxalacetate er en kraftig hemmer, mens succinatet favoriserer dissosiasjonen av oksalacetat, og fungerer som en aktivator.
Impulcinatmangel dehydrogenase
Den suksinatmangel dehydrogenase er en anomali eller mitokondriell respirasjonskjedeforstyrrelse. Denne mangelen er forårsaket av SDHA (eller SDHAF1), SDHB, SDHC og SDHD -mutasjoner.
Ulike undersøkelser har vist homozygote og heterozygote mutasjoner i disse genene, spesielt SDHA. Mutasjonene av disse genene forårsaker aminosyresubstitusjoner i proteinet (i noen av SDHA, B, C eller D), eller mislykkes som kodifiserer unormalt korte proteiner.
Derfor gir aminosyresubstitusjoner og unormalt kort proteinkoding, noe. Dette er hva forskere kaller som mitokondriell respirasjonskjedeforstyrrelse.
Denne lidelsen kan uttrykkes i mennesket fenotypisk på mange måter. De mest kjente er: mangel eller mangel på språklig utvikling, spastisk firedobling, muskel ufrivillige sammentrekninger (dystoni), muskelsvakhet og myokardiopati, blant andre relaterte problemer.
Noen pasienter med dehydrogenase for suksinatmangel kan manifestere Leights sykdom eller Kearns-saire syndrom.
Hvordan oppdages dehydrogenert succinatmangel?
Visse studier antyder bruk av kvalitative tester og analyser, samt kvantitativ, enzymatisk, luftveiskjede biokjemisk analyse. Andre antyder derimot fullstendig utvidelse av polymerasekjedereaksjonen (PCR) til eksonene til underenhetene som studeres og deretter, den respektive sekvensering.
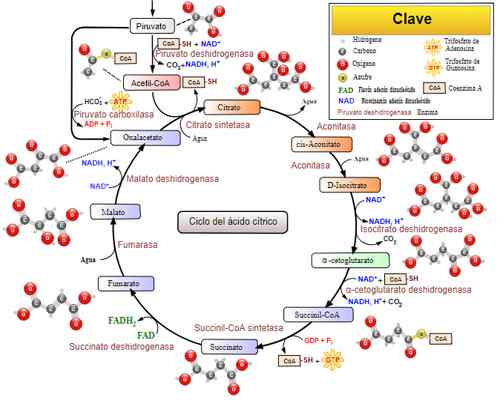 Tricarboxylic Acid Cycle (Krebs Cycle). Tatt og redigert fra: Narayanese, Wikiuserpedia, Yassinemraber, Totobaggins (oversatt til spansk av Alejandro Porto) [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)].
Tricarboxylic Acid Cycle (Krebs Cycle). Tatt og redigert fra: Narayanese, Wikiuserpedia, Yassinemraber, Totobaggins (oversatt til spansk av Alejandro Porto) [CC BY-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)]. Relaterte sykdommer
Det er mange fenotypiske uttrykk produsert av mitokondrielle respirasjonskjedesykdommer, på grunn av suksinatmangel dehydrogenase. Men når det handler om syndromer eller sykdommer, er det snakk om følgende.
Kan tjene deg: de 8 viktigste biogeokjemiske syklusene (beskrivelse)Leight syndrom
Det er en progressiv nevrologisk sykdom, assosiert med mutasjoner i det nukleære genomet (i dette tilfellet dehydrogenase succinat), som påvirker pyruvat-dishydrogenase-komplekset til den oksidative fosforyleringsveien.
Symptomene vises før det første året av individet, men i atypiske tilfeller har de første symptomene i ungdomsårene blitt observert.
Blant de mest observerte symptomene er: hypotoni med tap av kefalisk kontroll, ufrivillige bevegelser, tilbakevendende oppkast, luftveisproblemer, manglende evne til å flytte de okulære blodcellene, pyramide og ekstrapiramidale tegn blant andre blant andre blant andre. Anfall er ikke veldig vanlig.
Det er mulig at sykdommen kan oppdages i prenatal diagnoser. En spesifikk kur eller behandling er ikke kjent, men noen spesialister foreslår behandlinger med visse vitamin eller kofaktorer.
Gastrointestinal stroma tumor (GIST)
Vanligvis kalt GIST, er det en type gastrointestinal kanal -svulst, som generelt utvikler seg i områder som mage eller tynntarmen. Det antas at årsaken til disse skyldes en viss gruppe høyt spesialiserte celler som kalles ICC -celler eller mellomliggende celler i Cajal.
Andre hensyn til årsaken til GIST, er mutasjonene av visse typer gener, som ifølge noen forfattere forårsaker 90% av svulstene. Genene som er involvert er: Kit, PDGFRA -gener, Dehydrogenase Succinate (SDH) - Dårlig.
Succinatdehydrogenase (SDH) - dårlig, forekommer hovedsakelig hos unge kvinner, produserer magesvulster og produserer med relativ frekvens metastaser i lymfeknuter. En liten prosentandel forekommer hos barn, og i de fleste tilfeller skyldes det mangelen på uttrykk for SDHB -underenheten.
Kearns-Sayre syndrom
Det er blitt bestemt at noen pasienter med succinat dehydrogenase-mangler kan manifestere Kearns-Sayre-syndromet. Denne sykdommen er relatert til mitokondrielle lidelser, og er preget av fravær av bevegelse av okulære ballonger.
Andre kjennetegn ved denne sykdommen er pigmentrethet, døvhet, kardiomyopati og sentralnervesystemets forhold. Vanligvis blir disse symptomene observert før pasienten fyller 20 år. Ingen prenatal diagnose for denne tilstanden er kjent.
Heller ikke kur for denne sykdommen. Behandlingen er palliativ, det vil si at den bare fungerer for å redusere effekten av sykdommen, ikke kuren. På den annen side, selv om det avhenger av antall berørte organer og medisinsk behandling, er forventet levealder relativt normalt.
Referanser
- Ackrell, f. TIL., Kearney, e. B., & Singer, t. P. (1978). [47] Pattedyrs succinatdehydrogenase. I metoder i enzymologi (Vol. 53, s. 466-483). Akademisk presse.
- Brère, J. J., Favier, J., Ghouzzi, v. OG., Djoudi, f., Benit, s., Gimenez, a. P., & Rustin, P. (2005). Succinat dehydrogenase deficicy hos mennesker. Cellular and Molecular Life Sciences CMLS, 62 (19-20), 2317-2324.
- Cecchini, g., Schröder, i., Gunalus, r. P., & Maklashina, og. (2002). Succinat dehydrogenase og glansdyr redaktase fra Escherichia coli. Biochimica et Biophysica Acta (BBA) -Bioenergetics, 1553 (1-2), 140-157.
- Hatefi, og., & Davis, K. TIL. (1971). Succinat dehydrogenase. Yo. Rensing, molekylære egenskaper og understruktur. Biokjemi, 10 (13), 2509-2516.
- HEDERSTEDT, l. TIL. R. S., & Rutberg, L. TIL. R. S. (1981). Succinate dehydrogenase-a Comparative Review. Mikrobiologiske anmeldelser, 45 (4), 542.
- Nelson, d. L., Lehninger, a. L., & Cox, m. M. (2008). Lehninger prinsipper for biokjemi. Macmillan.
- Rutter, J., Winge, d. R., & Schiffman, J. D. (2010). Succinate dehydrogenase-ASEMBLEY, regulering og rolle i menneskelig diew. Mithochondrion, 10 (4), 393-401.
- « Essensielle fettsyrerfunksjoner, betydning, nomenklatur, eksempler
- Regel T -funksjoner, slik at det tjener, eksempler »