

Tyrosinegenskaper, struktur, funksjoner, fordeler
- 4567
- 39
- Anders Mathisen
De Tyrosin (Tyr, Y) er en av de 22 aminosyrene som utgjør proteinene til alle celler som er til stede i levende vesener. I motsetning til andre aminosyrer som valin, treonin, tryptofan, leucin, lysin og andre, er tyrosin en "betinget" i det vesentlige essensielt aminosyre.
Navnet "tyrosin" stammer fra det greske ordet "Skyting", som betyr ost, fordi denne aminosyren først ble oppdaget i denne maten. Begrepet ble myntet i 1846 av Liebig, som blandet ost med kaliumhydroksyd og oppnådde en ukjent forbindelse, neppe oppløselig i vann.
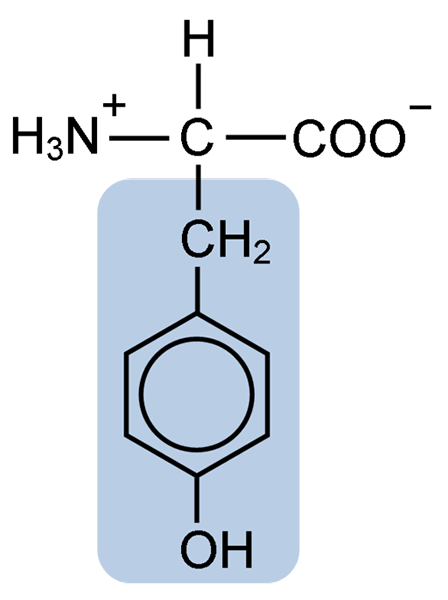
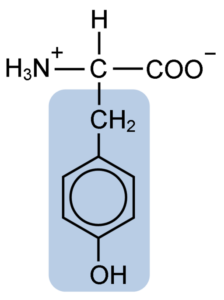 Kjemisk struktur av tyrosinaminosyren (kilde: clavecin [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons)
Kjemisk struktur av tyrosinaminosyren (kilde: clavecin [CC BY-SA 4.0 (https: // creativecommons.Org/lisenser/by-SA/4.0)] via Wikimedia Commons) Etter den første beskrivelsen, er andre forskere som Warren de La Rue og Hinterberger oppnådd fra henholdsvis kokoide insekter og hornproteiner. Dets separasjon fra hydrolyse av andre proteiner med saltsyre ble beskrevet i 1901 av Mörner.
Generelt oppnås denne aminosyren hos pattedyr takket være hydroksylering av fenylalanin, selv om den også blir absorbert i tarmen fra proteinene som konsumeres med mat.
Tyrosin har flere funksjoner i menneskekroppen, og blant disse er de mest relevante kanskje underlaget for produksjon av nevrotransmittere og hormoner som adrenalin og skjoldbruskhormon.
[TOC]
Kjennetegn
Tyrosin veier omtrent 180 g/mol og dens R- eller sidekjedegruppe har en 10 -dissosiasjonskonstant 10.07. Den relative overflod i celleproteiner overstiger ikke 4%, men har flere essensielle funksjoner for menneskelig fysiologi.
Denne aminosyren tilhører gruppen av aromatiske aminosyrer, der fenylalanin og tryptofan også er. Medlemmene av denne gruppen har aromatiske ringer i sine r r eller sidekjeder, og vanligvis er de hydrofobe eller apolare aminosyrer.
I likhet med tryptofan absorberer tyrosin ultrafiolett lys og er et av aminosyreavfallet som er ansvarlig for absorbansen av lys til 280 nm av mange proteiner, så det er nyttig for sin karakterisering.
Det regnes som en essensiell "betinget" aminosyre siden biosyntesen hos mennesket avhenger av fenylalanin, en essensiell aminosyre. Hvis kroppen tilfredsstiller sine daglige krav til fenylalanin, kan tyrosin syntetiseres uten problemer og er ikke en begrensende aminosyre.
Imidlertid, hvis kostholdet mangler fenylalanin, vil kroppen ikke bare ha en ubalanse av denne aminosyren, men også av tyrosin. Det er også viktig å påpeke at reaksjonen av tyrosinsyntese fra fenylalanin ikke er reversibel, så tyrosin kan ikke dekke de cellulære behovene til fenylalanin.
Tyrosin tilhører også aminosyregruppen med doble funksjoner i produksjonen av glukogene og ketogene metabolske mellommenn, som deltar i syntesen av glukose for hjernen og i dannelsen av ketonlegemer i leveren.
Struktur
Som resten av aminosyrene, tyrosinet eller β-parahydroksyfenyl-a-amino-propionsyre, er det en a-aminosyre som har et sentralt karbonatom, kalt karbon α og som er chiral, fordi det er knyttet til fire forskjellige substituenteatomer eller molekyler.
Dette quirale karbonet er knyttet til to karakteristiske grupper av aminosyrer: en aminogruppe (NH2) og en karboksylgruppe (COOH). Han deler også en av sine bindinger med et hydrogenatom, og den gjenværende bindingen er okkupert av R eller sidekjeden til hver aminosyre.
Når det.
Det kan tjene deg: BaroreceptorsFunksjoner
Tyrosine er en grunnleggende komponent av mange proteiner med stort mangfold av biologiske aktiviteter og funksjoner.
Hos mennesker og andre pattedyr blir denne aminosyren utnyttet i nerve- og nyrevev for syntese av dopamin, adrenalin og noradrenalin, tre nevrotransmittere relaterte katekolaminergika relatert til største betydning for kroppsfunksjon.
Det er også viktig for syntese av ultrafiolett strålingsbeskyttere (UV) som melanin; av noen smertestillende midler som endorfiner og antioksidantmolekyler som vitamin E.
Tilsvarende tjener denne aminosyren for syntese av tiramin-, oktopamin- og skjoldbruskhormoner gjennom jodorganisasjonen i tyrosinresten av tyoglobulin.
Tiramin er et vasoaktivt molekyl som finnes i menneskekroppen og oktopamin er et amin relatert til noreprenalin.
Alle disse funksjonene til tyrosin er mulig takket være at det oppnår fra proteinene i kostholdet eller ved hydroksylering av fenylalanin med leveren som hovedsystem for systemisk tilførsel av nevnte aminosyre.
Funksjoner i planter
Tyrosin og noen av mellommennene som ble generert under deres biosyntese mater de biosyntetiske rutene til metabolitter som er spesialisert i forsvar, i tiltrekningen av pollinatorer, i elektronisk transport og strukturell støtte.
Biosyntese
Hos mennesket oppnås tyrosin fra kostholdet eller syntetiseres i et enkelt trinn av leverceller fra fenylalanin, en essensiell aminosyre, gjennom reaksjonen katalysert av fenylalaninhydroksylase enzymatisk kompleks.
Dette komplekset har oksygenase -aktivitet og er bare til stede i leveren av mennesker eller andre pattedyr. Tyrosinsyntese -reaksjon innebærer deretter overføring av et oksygenatom til posisjonen til av den aromatiske ringen av fenylalanin.
Denne reaksjonen oppstår samtidig som et vannmolekyl dannes på grunn av reduksjon av et annet molekylært oksygenatom og den reduserende effekten blir bidratt direkte av en NADPH konjugert med et tetrahydropterinmolekyl, som ligner på folsyre.
Biosyntese i planter
I planter syntetiseres tyrosin av novo Nedstrøms for "Shikimato" -ruten, som mater andre biosyntetiske ruter av andre aromatiske aminosyrer som fenylalanin og tryptofan.
I disse organismer er syntesen basert på en forbindelse kjent som "corismato", som er sluttproduktet av Shikimato -ruten, og i tillegg den vanlige forløperen for alle aromatiske aminosyrer, visse vitaminer og plantehormoner.
Korismen omdannes til forord ved den katalytiske virkningen av mutasa -korismen enzymet, og dette er det første "engasjerte" trinnet i syntesen av tyrosin og fenylalanin i planter.
Predenate blir omdannet til tyrosin ved en oksidativ dekarboksylering og en transaminering, som kan oppstå i hvilken som helst rekkefølge.
I en av de biosyntetiske rutene kan disse trinnene katalyseres av spesifikke enzymer kjent som prefenatspesifikk dehydrogenase (PDH) (som konverterer forord henholdsvis.
En annen syntese av tyrosin fra forord innebærer transaminering av vidunderet til en ikke-proteinogen aminosyre kalt L-arogenat, katalysert av enzymet preenataminotransferase.
L-arogenatet blir deretter utsatt for oksidativ dekarboksylering for å danne tyroksin, en reaksjon regissert av et enzym tyrosin arogenat-spesifikk dehydrogenase, også kjent som ADH.
Planter bruker fortrinnsvis ruten til arenatet, mens de fleste mikrober syntetiserer tyrosin fra HPP avledet fra Predenesate.
Regulering
Som det er sant for de fleste av de biosyntetiske rutene med aminosyrer, har planter et strengt system for regulering av syntese av aromatiske aminosyrer, inkludert tyrosin.
Kan tjene deg: deilig monster: Kjennetegn, habitat, omsorg, egenskaperI disse organismer skjer regulering på mange nivåer, siden mekanismene som kontrollerer Shikimato -ruten også kontrollerer produksjonen av tyrosin, en rute som det også er deres egne reguleringsmekanismer.
Imidlertid er tyrosinbehov og derfor stivheten i reguleringen av deres biosyntese, spesifikk for hver planteart.
Nedbrytning
Nedbrytning eller katabolisme av tyrosin resulterer i dannelse av fumarat og acetoacetat. Det første trinnet i denne ruten består i omdannelsen av aminosyren til 4-hydroksyfenylpypat ved et cytosolisk enzym kjent som aminotransferase-tyrosin.
Denne aminosyren kan også transamineres i mitokondriene til hepatocytter ved et aminotransferase aspartatenzym, selv om dette enzymet ikke er veldig viktig under normale fysiologiske forhold.
Gjennom nedbrytning av tyrosin kan succinyl-acetoacetat forekomme, som kan dekarboksyleres til succinyl-acetat. Succinyl-acetat er den kraftigste enzyminhibitoren som er ansvarlig for syntesen av hemokruppen, 5-aminolevulinsyren dehydratase.
Adrenalin og noradrenalinsyntese
Som nevnt er tyrosin et av hovedunderlagene for syntese av to veldig viktige nevrotransmittere for menneskekroppen: adrenalin og noradrenalinin.
Dette brukes opprinnelig av et enzym kjent som hydroksylase -tyrosin, i stand til å tilsette en ekstra hydroksylgruppe til den aromatiske ringen til RI tyrosin R -gruppen, som danner forbindelsen kjent som DOPA.
DOPA gir opphav til dopamin når den først er enzymatisk behandlet av et enzym DOPA -dekboksilas.
Dopamin blir deretter omdannet til noreprenalin ved virkningen av β -oksidase -dopaminenzymet, som katalyserer tilsetningen av en hydroksylgruppe til -ch som var en del av gruppen R av tyrosin og som fungerte som en "bro" mellom den aromatiske ringen og karbon α.
Adrenalin stammer fra noradrenalin ved virkning av fenytanolamin N-Metyltransferase, som er ansvarlig for S-adenosyl-metioninavhengig overføring av en metylgruppe (-Ch3) til den frie aminogruppen av norepinefrin.
Mat rik på tyrosin
Som nevnt ovenfor er tyrosin en essensiell "betinget" aminosyre, siden den syntetiseres i menneskekroppen ved hydroksylering av fenylalanin, en essensiell aminosyre.
Derfor, hvis inntak av fenylalanin dekker kroppskrav, er tyrosin ikke en begrensende faktor for normal funksjon av celler. Tyrosin er imidlertid også anskaffet fra proteiner som konsumeres med daglig mat.
Noen studier rapporterer at minimum daglig inntak av både tyrosin og fenylalanin bør være mellom 25 og 30 mg per kilo vekt, så en gjennomsnittlig person bør konsumere mer eller mindre 875 mg tyrosin per dag.
Mat med høyere tyrosininnhold er ost og soyabønner. Blant disse er også storfekjøtt, lam, svinekjøtt, kylling og fisk.
Noen frø og nøtter som nøtter gir også betydelige bidrag fra denne aminosyren, samt egg, meieriprodukter, korn og frokostblandinger.
Fordelene med inntaket ditt
Tyrosin blir ofte konsumert i form av kostholds- eller kosttilskudd, spesielt for behandling av patologi kjent som fenylcetonuria, påført av pasienter som ikke er i stand til å behandle fenylalanin på riktig måte og produserer derfor ikke tyrosin.
Det kan tjene deg: forholdsfunksjonDet antas at en økning i mengden tyrosin som konsumeres per dag kan forbedre kognitive funksjoner relatert til læring, hukommelse og varselstilstand under stressende forhold, siden den.
Noen mennesker tar tirosin -rike tabletter for å holde seg våken om dagen hvis de har mistet en betydelig mengde søvntider.
Siden denne aminosyren deltar i dannelsen av skjoldbruskkjertelhormon, kan forbruket ha positive effekter på systemisk metabolsk regulering.
Mangelforstyrrelser
Albinisme og alcaptonuria er to patologier relatert til tyrosinmetabolisme. Den første tilstanden er relatert til mangelfull melaninsyntese fra tyrosin, og den andre har å gjøre med defekter i tyrosin -nedbrytning.
Albinisme er preget av mangelen på pigmentering i huden, det vil si at pasientene som lider av det har hvitt hår og rosa hud, fordi melanin er pigmentet som er ansvarlig for å gi farge til disse strukturene.
Denne patologien har å gjøre med en mangel i den spesifikke tyrosinasen til melanocytter, som er ansvarlig for å konvertere tyrosin til Dopa-Quinone, en mellomledd i syntesen av melanin.
Symptomene på Alcaptonuria er tydelige som en overdrevet urinpigmentering (mørk) og sen utvikling leddgikt.
Andre metabolske patologier
I tillegg er det andre lidelser relatert til tyrosinmetabolisme, blant dem:
- Arvelig tyrosinemi av type I: karakterisert av progressiv leverdegenerasjon og nyrefunksjon
- Arvelig tyrosinemi av type II eller Richner-Hanhart-syndrom: som er tydelig som en keratitt og ampulære lesjoner i håndflatene og sålene på føttene
- Tyrosinemia av type III: som kan være asymptomatisk eller til stede som mentale forsinkelser
- "Hawkinsinuria": preget av metabolsk acidose i barndommen og manglende evne til å blomstre i vekst
Det er også andre medfødte defekter i metrosinmetabolismen som har å gjøre med mangler i enzymene som.
Referanser
- Aders plimmer, r. (1908). Proteinerens kjemiske konstitusjon. jeg dro. London, Storbritannia: Longmans, Green og CO.
- Chakrapani, a., Gissen, p., & McKiernan, P. (2012). Forstyrrelser av tyrosinmetabolisme. I Medfødt metabolsk ønske: Diagnose og behandling (s. 265-276).
- Kretchmer, n., Levine, s., McNamara, h., & Barnett, h. (1956). Visse aspekter ved tyrosinmetabolisme hos de unge. Yo. Utviklingen av tyrosinoksideringssystemet i menneskelig lever. Journal of Clinical Investigation, 35(10), 1089-1093.
- Du, f., Zannoni, v., Laster, l., & Seegmiller, e. (1958). Naturen til defekten ved tyrosinmetabolisme i Alkaptonuria. Journal of Biological Chemistry, 230, 251-260.
- Murray, r., Bender, d., Botham, k., Kennelly, p., Rodwell, v., & Weil, P. (2009). Harpers illustrerte biokjemi (28. utg.). McGraw-Hill Medical.
- Nelson, d. L., & Cox, m. M. (2009). Lehninger prinsipper for biokjemi. Omega -utgaver (5. utg.).
- Schenck, c. TIL., & Maeda, h. TIL. (2018). Tyrosinbiosyntese, metabolisme og katabolisme i planter. Fytokjemi, 149, 82-102.
- Slominski, a., Zmijewski, m. TIL., & Paweley, J. (2012). L-tyrosin og L-dihydroksyfenylalanin som hormonlignende regulatorer av melanocyttfunksjoner. Pigmentcelle og melanomforskning, 25(1), 14-27.
- Van de, g. (2018). Helselinje. Hentet 16. september 2019, fra www.Healthline.com
- MD Web. (n.d.). Hentet 15. september 2019, fra www.Webmd.com
- Whitbread, d. (2019). Matdataene mine. Hentet 15. september 2019, fra www.Myfooddata.com