

Tre egenskaper og funksjoner i organismen
- 1801
- 11
- Jonathan Moe
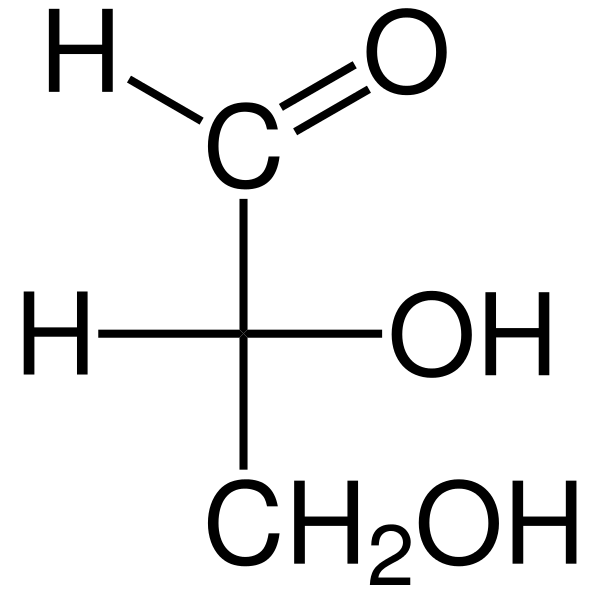
De Triosas De er tre -karbon monosakkarider hvis empiriske kjemiske formel er c3H6ENTEN6. Det er to triosas: glyseraldehyd. Triosasene er viktige i metabolismen fordi de forbinder tre metabolske veier: glykolyse, glukoneogenese og pentosefosfatbanen.
Under fotosyntesen er Calvin-syklusen en kilde til triosas som tjener til fruktose-6-fosfatbiosyntese. Dette sukkeret, på fosforylert måte, omdannes med enzymatisk katalyserte trinn til reserve eller strukturelle polysakkarider.
Kilde: Wesalius [Public Domain] Triosasene deltar i biosyntesen av lipider som er en del av cellemembraner og adipocytter.
[TOC]
Kjennetegn
Glyceraldehyd aldosa har et quiralt karbonatom, og derfor har det to enantiomeros, L-glyceraldehyd og D-glyceraldehyd. Begge enantiomerer D og L har forskjellige kjemiske og fysiske egenskaper.
D-glyseraldehydet ødela planet til det polariserte lyset til høyre (+) og har en rotasjon [α] D, ved 25 ° C, fra +8.7 °, mens L-glyseraldehydet ødelagte planet til det polariserte lyset på venstre (-) og har en rotasjon [α] D, ved 25 ° C, fra -8.7 °.
Det chirale karbonet i glyseraldehyd er karbon 2 (C-2), som er en sekundær alkohol. Fischers projeksjon representerer hydroksylgruppen (-OH) til D-glyceraldehyd til høyre og oh-gruppen i venstre l-glyce.
Dihydroxyacetone mangler chirale karbonatomer og har ikke enantiomere former. Tilsetningen av en hydroksymetylengruppe (-CHOH) til glyseraldehyd eller dihydroksyaceton tillater opprettelse av et nytt chiralsenter. Følgelig er sukker en tetrosa fordi den har fire karbonatomer.
Tillegg av en gruppe -chah til Terosa skaper et nytt chiralsenter. Dannet sukker er en pentose. Grupper -chah kan fortsette å legges til maksimalt ti karbonatomer.
Kan tjene deg: fylogeniFunksjoner i organismen
Triosasene som mellommenn i glykolyse, glukoneogenese og pentosa fosfatbane
Glykolyse består av brudd på glukosemolekylet i to pyruvatmolekyler for å produsere energi. Denne ruten innebærer to faser: 1) forberedende fase, eller energiforbruk; 2) Fase av energiproduksjon. Den første er den som produserer triosas.
I den første fasen økes glukosefrie energiinnholdet ved dannelse av fosfoésters. I denne fasen er adenosin -triffosfatet (ATP) fosfatdonoren. Denne fasen kulminerer med konvertering av fruktosefosfoéster 1,6-bifosfat (F1,6bp) i to triosas fosfat, glyseraldehyd 3-fosfat (GA3P) og dihydroksyaceton-fosfat (DHAP).
Glukoneogenese er glukosebiosyntese fra pyruvat og andre mellommenn. Bruk alle glykolyse -enzymer som katalyserer reaksjoner hvis biokjemisk standard energivariasjon er i likevekt (Δgº '~ 0). På grunn av dette har glykolyse og glukoneogenese vanlige formidlere, inkludert GA3P og DHAP.
Pentosefosfatbanen består av to trinn: en oksidativ fase av glukose-6-fosfat og en annen dannelse av NADPH og Ribosa-5-fosfat. I den andre fasen omdannes 5-fosfat ribose til glykolyse-mellommenn, F1.6bp og Ga3p.
Triosas og Calvin -syklusen
Fotosyntesen er delt inn i to stadier. I de første oppstår lysavhengige reaksjoner som produserer NADPH og ATP. Disse stoffene brukes i det andre, der det er fiksering av karbondioksid og heksosedannelse fra triosas gjennom en sti kjent som Calvin Cycle.
I Calvin-syklusen katalyserer den 1,5-bifariske oksygenase (Rubisco) ribuloseenzymet den kovalente foreningen av CO2 Til pentose ribulosa 1,5-bifosfat og bryter den ustabile mellomledningen av seks karbonatomer i to molekyler av tre karbonatomer: 3-fosfoglyseratet.
Kan tjene deg: oligosakkarider: egenskaper, sammensetning, funksjoner, typerGjennom enzymatiske reaksjoner som inkluderer fosforylering og reduksjon av 3-fosfoglyserat, ved bruk av ATP og NADP, forekommer Ga3p. Denne metabolitten omdannes til 1,6-bifosfat fruktose (F1.6bp) av en metabolsk vei som ligner på glukoneogenese.
Ved handling av en fosfatase blir F1.6bp omdannet til fruktose-6-fosfat. Deretter produserer en isomease fosfhexose glukose 6-fosfat (GLC6P). Til slutt konverterer en epicherase GLC6P til glukose 1-fosfat, som tjener for stivelsesbiosyntese.
Triosas og lipider av biologiske og adipocyttmembraner
GA3P og DHAP kan danne fosfatglyserol som er en nødvendig metabolitt for biosyntesen av triacylglyseroler og glyserolipider. Dette er fordi begge triosasfosfat kan interkonverteres av en reaksjon katalysert av triosa -isomerasefosfat, som holder begge triosaer i likevekt.
Glyserol-fosfatenzymet dehydrogenase katalyserer en oksid-reduksjonsreaksjon, der NADH donerer et par elektroner til DHAP for å danne 3-fosfatglyserol og NAD+. L-glyserol 3-fosfat er en del av skjelettet til fosfolipider som er strukturell del av biologiske membraner.
Glyserol er proquiral, den mangler asymmetriske karbonatomer, men når en av de to primære alkoholer danner en fosfoester, kan den riktig kalles L-glycerol 3-fosfat, eller D-glyserol 3-fosfat.
Glyceofosfolipider kalles også fosfoglyserider, og blir navngitt som fosfatidinsyre avledet. Fosfoglyserider kan danne fosfoacylglyceroler ved å danne esterbindinger med to fettsyrer. I dette tilfellet er det resulterende produktet 1,2-fosfodiacylglycerol, som er en viktig komponent i membranene.
En glyceofosfase katalyserer hydrolysen av 3-fosfatglyserolfosfatgruppen, og produserer glyserol mer fosfat. Glyserol kan tjene som en startmetabolitt for biosyntese av triacylglycerider, som er vanlig i adipocytter.
Kan tjene deg: Coagulase Test: Foundation, Procedure and UsesTriosas og membraner i arkeobakteriene
I likhet med eubakterier og eukaryoter, dannes 3-fosfatglyserol fra triosas fosfat (GA3P og DHAP). Imidlertid er det forskjeller: det første er at 3-fosfatglyserol i arkeobakterier membraner er L-konfigurasjon, mens det i eubakterier og eukaryote membraner er konfigurasjon d.
En annen forskjell er at arkeobakterier membraner danner esterkoblinger med to lange hydrokarbonkjeder av isoprenoidgrupper, mens i eubakterier og eukaryoter er glyserol danner esterbindinger (1,2-diacilglycerol) med to hydrokarbonkjeder av fettsyrer.
En tredje forskjell er at erstatninger i fosfat og 3-fosfatgysfatgysfatgysfatgysfat glyserol er forskjellige fra eubakterier og eukaryoter og eukaryoter. For eksempel er fosfatgruppen knyttet til disakkarid α-Glucopiranso- (1®2)-β-Galatofuranosa.
Referanser
- Cui, s. W. 2005. Matkarbohydrater: Kjemi, fysiske egenskaper og applikasjoner. CRC Press, Boca Raton.
- Cock, s., Mäkinen, K, Honkala, og., Saag, m., Kennepohl, e., Eapen, a. 2016. Erythritol er mer effektiv enn xylitol og sorbitol i å håndtere orale helseendepunkter. International Journal of Dentistry.
- Nelson, d. L., Cox, m. M. 2017. Lehninger prinsipper for biokjemi. W. H. Freeman, New York.
- Sinnott, m. L. 2007. Karbohydratkjemi og biokjemi -struktur og mekanisme. Royal Society of Chemistry, Cambridge.
- Stick, r. V., Williams, s. J. 2009. Karbohydrater: De essensielle molekylene i livet. Elsevier, Amsterdam.
- Voet, d., Voet, J. G., Pratt, c. W. 2008. Grunnleggende om biokjemi - Liv på molekylært nivå. Wiley, Hoboken.
- « Aldoseegenskaper, typer, antall karbonatomer
- Fluoksetin (prozac) virkningsmekanisme og bivirkninger »