

Uracilstruktur, funksjoner, egenskaper, syntese
- 2271
- 244
- Theodor Anders Hopland
Han Uracil Det er en pyrimidin nitrogenbase, som finnes i ribonukleinsyre (RNA). Dette er en av egenskapene som skiller RNA fra deoksyribonukleinsyre (DNA), siden sistnevnte har Timina i stedet for uracil. Både stoffer, Uracil og Timina, er bare forskjellige at den andre har en metylgruppe.
Fra det evolusjonære synspunktet har det blitt foreslått at RNA var det første molekylet som lagret genetisk informasjon og fungerte som en katalysator i cellene, før DNA og enzymer. På grunn av dette antas det at Uracil hadde en nøkkelrolle i utviklingen av livet.
Kilde: Kemikungen [Public Domain] Hos levende vesener finnes ikke uracil fritt, men ofte monofosfatnukleotider (UMP), difosfat (UDP) og Trphosphate (UTP). Disse uracil -nukleotidene har forskjellige funksjoner, for eksempel RNA -biosyntese og glykogen, isomer interkonvertering av sukker og syntase -syntetaminregulering.
[TOC]
Struktur og egenskaper
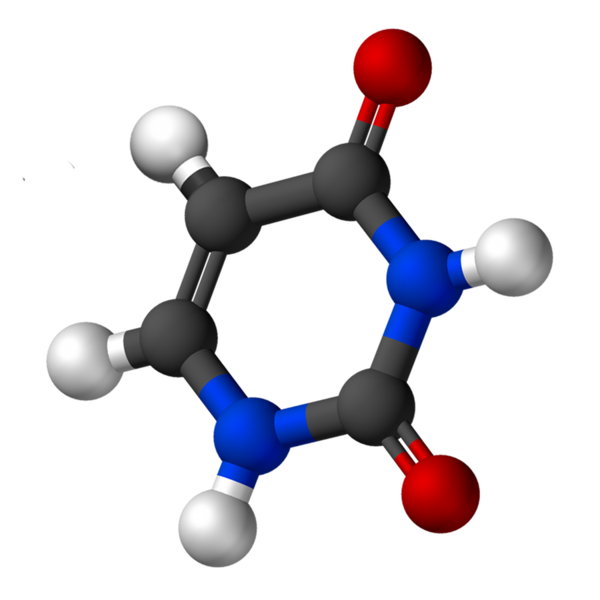
Uracil, kalt 2,4-dioxipiridin, har den empiriske formelen C4H4N2ENTEN2, hvis molekylvekt er 112,09 g/mol, og blir renset som et hvitt pulver.
Strukturen til uridin er en heterocyklisk ring med fire karbonatomer og to nitrogen, med doble alternative bindinger. Det er plan.
Den har en 50 mg/ml løselighet, 25 ºC, i 1 m natriumhydroksyd, og en PKA mellom 7.9 og 8.2. Bølgelengden der den maksimale absorbansen oppstår (ʎMaks) Det er mellom 258 og 260 nm.
Biosyntese
Det er en vanlig vei for biosyntese av pyrimidin -nukleotider (uracil og cytokin). Det første trinnet er biosyntesen av karbamoilfosfat fra CO2 og NH4+, som katalyseres av karbamoilsyntetase fosfat.
Pyrimidin er bygget av karboilfosfat og aspartat. Begge stoffene reagerer og danner N-karbamoilaspartato, reaksjon katalysert av Transcabamoilasa aspartat (ATCASA). Stengingen av pyrimidinringen kan dehydreres med dihydrootase, og produserer L-dihydrotatatoat.
L-dihydrotoatet oksideres og blir omgjort til orotat; Elektronakseptoren er NAD+. Det er en reaksjon katalysert av dihydroorotatdehydrogenase. Neste trinn består i overføringen av fosforibosylgruppen, fra fosforibosylpyrofosfat (PRPP), til orotat. Gradidilato form (WPO) og uorganisk piroffosfat (PPI), katalysert av fosforibosylorotatoverføringen.
Det siste trinnet består av dekarboksylering av pyrimidinringen til gruppen (WPO). Uridilaato form (uridin-5'-monofosfat, UMP), som er katalysert av en dekarboksylase.
Deretter overføres en fosfatgruppe gjennom deltakelse av en kinase fra ATP til UMP, og danner UDP (uridin-5'-difosfat). Sistnevnte gjentas og danner UTP (uridin-5'-triffosfat).
Kan tjene deg: grener av biologi og hva studerer deBiosynteseforordning
I bakterier skjer reguleringen av pyrimidinbiosyntese gjennom negativ tilbakemelding, ved transcabamoilasa aspartatnivå (ATCASA).
Dette enzymet blir hemmet av CTP (Citidin-5'-Trophosphate), som er sluttproduktet av biosynteseveien til pyrimidin. ATCAS.
Hos dyr skjer reguleringen av pyrimidinbiosyntese gjennom negativ tilbakemelding, på nivå med to enzymer: 1) karbamoilsyntase II -fosfat, som blir hemmet av UTP og aktivert av ATP og PRPP; og 2) OMP -disarboxylase, som er hemmet av produktet av reaksjonen som hun selv katalyserer, UMP. Biosyntesehastigheten til OMP varierer med tilgjengeligheten av PRPP.
Funksjon i biosyntesen av RNA
Uracil er til stede i alle typer RNA, for eksempel Messenger RNA (MNA), overførings -RNA (ARNT) og ribosomal RNA (RNA). Biosyntesen av disse molekylene skjer gjennom en prosess som kalles transkripsjon.
Under transkripsjonen kopieres informasjonen i DNA i RNA gjennom et Pon -polymerase -RNA. Den omvendte prosessen, der informasjonen i RNA er kopiert i DNA, skjer i noen virus og planter gjennom omvendt transkriptase.
RNA -biosyntese trenger tryposfatnukleosider (NTP), nemlig: tryfosfat uridin (UTP), tryposfatcytidin (CTP), adenin tryposfat adenin (ATP) og tryphosphate guanin (GTP). Reaksjonen er:
(RNA)n avfall + NTP -> (RNA)N+1 Avfall + PPI
Uorganisk pyrofosfat (PPI) hydrolyse gir energi for biosyntesen av RNA.
Funksjon i sukkerbiosyntese
Sukkerestere er veldig vanlig i levende organismer. Noen av disse estere er difosfaene til nukleosidestere, for eksempel UDP-sukker, som er veldig rikelig i celler. UDP-sukker involverer biosyntesen av disakkarider, oligosakkarider og polysakkarider.
Hos planter skjer biosyntese av sukrose på to måter: en hoved- og sekundær.
Hovedruten består i overføringen av D-glukosen fra UDP-D-glukosa til D-Frucease for å danne sukrose og UDP. Sekundærveien inkluderer to trinn: Den starter med UDP-D-glukose og fruktose-6-fosfat og kulminerer med dannelse av sukrose og fosfat.
I brystkjertlene skjer laktosebiosyntese fra UDP-D-galaktose og glukose.
Kan tjene deg: grener av biokjemiHos planter utføres cellulosebiosyntese ved kontinuerlig kondensering av beta-d-glykosylavfall, fra UDP-glukose til den ikke-reduserende enden av den voksende polyglukosekjeden. Tilsvarende krever amylose og amylopektinbiosyntese.
Hos dyr brukes både UDP-glukose og ADP-glukose til glykogenbiosyntese. Tilsvarende krever kondroitinsulfatbiosyntese UDP-Xylose, UDP-Galactose og UDP-Glucuronato.
Funksjon i isomer interkonversjon av sukkerarter
Konvertering av galaktose til en formidler av glykolyse skjer gjennom leloirveien. Et av trinnene på denne ruten blir katalysert av UDP-galaktose-4-epimerase-enzymet, som letter interkonversjonen av UDP-galaktose til UDP-glukose.
Funksjon i biosyntese av glykoproteiner
Under biosyntesen av glykoproteiner går proteiner gjennom cis, medium og trans -sekker av Golgi -apparatet.
Hver av disse posene har et sett med enzymer som behandler glykoproteiner. Sukkermonomerer, som glukose og galaktose.
Nukleotidene-hexose blir transportert til Golgi-tanker med Antiporte. UDP-Galactose (UDP-Gal) og UDP-N-acetylgalactosamina (UDP-Galnac) går inn i tankene fra cytosol ved hjelp av UMP.
I Golgi -tanken hydrerer en fosfatase en fosfatgruppe av UDP og UMP og PI. UDP kommer fra reaksjonene katalysert av galaktosyltransferase og N-acetylgalaktosamiltransferase. UMP dannet av fosfatase tjener til å utveksle nukleotider-heksose.
Funksjon i reguleringen av syntase
En mekanisme for å regulere glutaminsyntase er den kovalente modifiseringen, som består av adenilering, som er inaktiv, og flanke, som aktivt den. Denne kovalente modifiseringen er reversibel og katalysert av adenyltransferase.
Aktiviteten til adenyltransferase moduleres av foreningen av PII -proteinet, som er regulert av en kovalent modifisering, uridinilering.
Både uridililering og forskyvningen utføres av uridililtransferase. I dette enzymet skyldes uridilasjonsaktiviteten glutamin og fosfat, og aktiveres av foreningen av alfa-zotoglutarat og ATP til PII.
Funksjon i RNA -utgave
Noen RNM redigeres før oversettelse. I noen eukaryote organismer, for eksempel Trypanosoma Brucei, Det er utgave av RNA for transkripsjonen av underenheten II -genet til cytokromoksidase. Dette skjer ved å sette inn uracil avfall, en reaksjon katalysert av terminalen uridiltransferase.
Kan tjene deg: krøll: komposisjon, deler, betydningEn guide RNA, komplementær til det redigerte produktet, fungerer som temperert for redigeringsprosessen. Baseparene dannet mellom det første transkriptet og guide-RNA innebærer basepar g = u som ikke er Watson-rekord og er vanlig i RNA.
UDP-glukosebiosyntese
Under fysiologiske forhold er glykogenbiosyntese fra glukose-1-fosfat termodynamisk umulig (positiv ΔG). På grunn av dette, før biosyntese, skjer aktivering av glukose-1-fosfat (G1P). Denne G1P og UTP kombinerer reaksjon for å danne glukosedifosfat uridin (UDP-glukose eller UDPG).
Reaksjonen katalyseres av pyrofosforyllasen til UDP-glukosen, og er som følger:
G1P + UTP -> UDP -GLUCOSA + 2PI.
Gibbs fri energivariasjon i dette trinnet er stort og negativt (-33,5 kJ/mol). Under oksygenreaksjon angriper G1P Alforo Alpha Phosphorus-atomet i UDP-glukose og uorganisk pyrofosfat (PPI). Deretter hydrolyseres PPI av en uorganiske pyrofosfater, hvis hydrolyseenergi er den som driver den generelle reaksjonen.
UDP-glukose er et "høy energi" -stoff. Det gjør det mulig å danne glykosidbindinger mellom glukoseresten og den økende polysakkaridkjeden. Det samme energiprinsippet gjelder reaksjonene der UDP-sukker deltar, for eksempel biosyntesen av disakkarider, oligosakkarider og glykoproteiner.
Uracil DNA glykosilasa
Det er DNA -lesjoner som forekommer spontant. En av disse lesjonene er sprontanen. I dette tilfellet skjer reparasjonen på grunn av den modifiserte DNA -basen ved hjelp av et enzym kalt uracil DNA -glykosilasa.
Enzymet uracil DNA-glykosilasa eliminerer det skadede cytokinet (uracil), og produserer en deoksyriboserest som mangler nitrogenbase, kalt AP-sted (apurinisk-apirimidinisk sted).
Deretter gjør Endonuklease AP-enzymet et kutt i fosfodiesterskjelettet på AP-stedet, og eliminerer sukker-fosfatresten. DNA -polymerase jeg gjenoppretter den skadede strengen.
Referanser
- Bohinski, r. 1991. Biokjemi. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokjemi. Redaksjonell reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, p., Baltimore, d., Darnell, J. 2003. Cellulær og molekylær biologi. Redaksjonell Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Mexico, Sāo Paulo.
- Nelson, d. L., Cox, m. M. 2008. Lehninger-prinsipper for biokjemi. W.H. Freeman, New York.
- Voet, d. og Voet, J. 2004. Biokjemi. John Wiley og sønner, USA.