

Cystring -klassifisering, trening, utvikling og segmentering
- 1981
- 488
- Anders Mathisen
Han Zygote Det er definert som cellen som er resultatet av fusjonen mellom to gameter, en kvinne og en maskulin. I følge den genetiske belastningen er zygoten diploid, noe som betyr at den inneholder den komplette genetiske belastningen til de aktuelle artene. Dette er fordi gametene som stammer fra det hver inneholder halvparten av artskromosomene.
Det er ofte kjent som egg og er strukturelt sammensatt av to pronomer, som kommer fra de to gametene som har sin opprinnelse. På samme måte er det omgitt av det pelukide området, som oppfyller en trippel funksjon: unngå at mellom en annen sæd, hold cellene som følge av de første divisjonene i zygoten sammen og forhindrer at implementeringen oppstår til zygoten når nettstedet idealet i livmor.
Zygote utvikling. Kilde: CNX OpenStax [CC av 4.0 (https: // creativecommons.Org/lisenser/av/4.0)] Cytoplasmaet til zygoten, så vel som organellene som er inneholdt i den, er av mors opprinnelse, siden de kommer fra eggløsningen.
[TOC]
Klassifisering
Zygoten er klassifisert i henhold til to kriterier: mengden vitellus og organisering av Vitellus.
-Typer zygote i henhold til mengden vitelo
Avhengig av mengden vitellus som zygoten har, kan dette være:
Oligolecito
Generelt sett er oligolecito zygote en som inneholder veldig lite vitelo. På samme måte er de i de fleste tilfeller redusert størrelse og kjernen har en sentral posisjon.
Et nysgjerrig faktum er at av denne typen egg de stammer fra, for det meste, larver som har fritt liv.
Den typen dyr som denne typen zygote blir verdsatt, er echinoderms, for eksempel pinnsvin og sjøstjerner; Noen ormer som platelmints og nematoder; bløtdyr som snegler og blekkspruter; og pattedyr som mennesket.
Mesolecito
Dette er et ord som består av to ord, "meso" som betyr halvparten, og "lecito" som betyr vitelo. Derfor er denne typen zygote en som har en moderat mengde vitellus. Tilsvarende er dette hovedsakelig lokalisert i en av zygotpolene.
Denne typen egg er representativ for noen virveldyr som amfibier, representert av frosker, padder og salamandere, blant andre.
Polilecito
Ordet polyilecito er dannet av ordene "poli", som betyr mye eller rikelig, og "lecito", som betyr vitelo. I denne forstand er polilecito zygoten en som inneholder en stor mengde vitel. I denne typen zygote er kjernen i en sentral posisjon av Vitle.
Polilecito zygoten er typisk for fugler, krypdyr og noen fisk som haier.
Typer zygote i henhold til organisering av Vitellus
I følge distribusjonen og organiseringen av Vitellus er zygoten klassifisert som:
Isolekito
Ordet Isolekito består av "ISO", som betyr det samme, og "Lecito", som betyr vitelo. På en slik måte at Zygote -typen er en der Vitellus presenterer en homogen distribusjon gjennom det tilgjengelige rommet.
Denne typen zygote er typisk for dyr som pattedyr og kråkeboller.
Telolecitos
I denne typen zygote er Vitell rikelig og okkuperer nesten all tilgjengelig plass. Cytoplasmaet er ganske liten og inneholder kjernen.
Denne zygoten er representativ for arter av fisk, fugler og krypdyr.
Sentroleter
Som det skal utledes med navn, er Vitle i denne typen egg i en sentral posisjon. På samme måte er kjernen lokalisert i midten av det vitelløse. Denne zygoten er preget av å være oval form.
Denne typen zygote er typisk for medlemmene i gruppen av leddyr, for eksempel arachnider og insekter.
Zygotedannelse
Zygoten er cellen som dannes umiddelbart etter befruktningsprosessen oppstår.
Befruktning
Befruktning er prosessen der kvinnelige og mannlige gameter er sammen med. Hos mennesker er den kvinnelige zygoten kjent som eggløsningen og den mannlige zygoten kalles sæd.
Det kan tjene deg: Bioelements: Klassifisering (Primær og sekundær)Tilsvarende er ikke befruktning en enkel og enkel prosess, men består av en serie stadier, hver veldig viktig, nemlig:
Kontakt og penetrering i den utstrålte kronen
Når sædcelleren etablerer den første kontakten med eggløsningen, gjør den det i det så -kallede pelukideområdet. Denne første kontakten er av transcendental betydning, siden den tjener for hver gamete å gjenkjenne den andre, og avgjøre om de tilhører samme art.
På samme måte er sædcellene i stand til å krysse et lag med celler som omgir eggløsningen, og som som helhet er kjent som utstrålt krone.
For å kunne krysse det laget av celler, utskiller sæd et enzymatisk stoff som kalles hyaluronidase som hjelper deg i prosessen. Et annet element som gjør at sæd kan trenge gjennom dette ytre laget av egget er den hektiske bevegelsen av halen.
Introduksjon til Pelucida -området
Når sædcellen har krysset den utstrålte kronen, står sædene overfor en annen hindring for å trenge gjennom eggløsningen: det pelukide området. Dette er ikke noe mer enn det ytre laget som omgir eggløsningen. Det består hovedsakelig av glykoproteiner.
Når sædhodet kommer i kontakt med pelukidområdet, utløses en reaksjon kjent som akrosomisk reaksjon. Dette består av frigjøring, av sæd, av enzymer som er kjent som spermiolisiner som helhet. Disse enzymene er lagret i et rom med sædhodet kjent som Acrosoma.
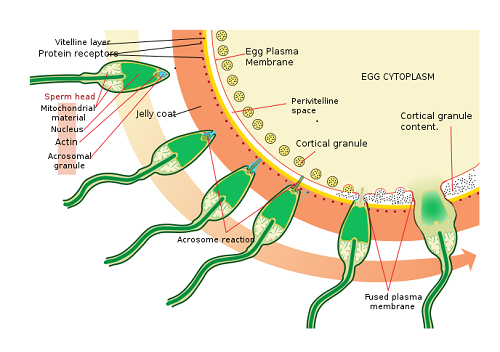 Akrosomisk reaksjon. Kilde: Ladyofhats. [Offentlig domene]
Akrosomisk reaksjon. Kilde: Ladyofhats. [Offentlig domene] Spermiolisinene er hydrolytiske enzymer som har som hovedfunksjon nedbrytningen av pelukidområdet, for endelig å trenge gjennom eggløsningen.
Når den akrosomiske reaksjonen begynner, utløses også en serie strukturelle forandringer på nivået av membranen.
Membranfusjon
Neste trinn i befruktningsprosessen er fusjon av membranene til de to gametene, det vil si eggløsningen og sædcellene.
Under denne prosessen skjer en serie transformasjoner i eggløsningen som tillater inntreden av en sæd og unngå inntreden av all annen sæd som omgir den.
For det første dannes en ledning kjent som en befruktningskegle, som sæd- og eggløsemembranene kommer i direkte kontakt, som ender ved å slå sammen.
Samtidig til dette skjer en ionemobilisering som kalsium (CA på eggløsemembranenivå (CA (Ca+2), Hydrogen (h+) og natrium (Na+), som genererer den så -kalt depolarisering av membranen. Dette betyr at polariteten som normalt har.
Tilsvarende er det under eggløsemembranen strukturer som kalles kortikale granuler, som frigjør innholdet til rommet rundt eggløsningen. Med dette er det som oppnås å forhindre overholdelse av sæd fra eggløsningen, slik at de ikke kan nærme seg dette.
Fusjon av eggløsekjerner og sæd
For endelig å danne zygoten, er det nødvendig at sæd- og eggløsningene blir sammenføyd.
Det er verdt å huske at gameter bare inneholder halvparten av antall artskromosomer. Når det gjelder mennesket, er det 23 kromosomer; Dette er grunnen til at de to kjernene må slås sammen for å danne en diploid celle, med den fullstendige genetiske belastningen til arten.
Når sædcellen kommer inn i eggløsningen, er DNAet doblet, så vel som DNA fra eggløsningen pronucleus. Deretter finnes begge pronucelos ved siden av den andre.
Umiddelbart går membranene som skiller både i oppløsning, og på denne måten kan kromosomene som var inneholdt i hver sammen komme sammen med sin motpart.
Men alt slutter ikke her. Kromosomer er lokalisert i ekvatorialstangen til cellen (zygote) for å starte den første av mange mitotiske divisjoner i segmenteringsprosessen.
Kan tjene deg: apolipoprotein e: egenskaper, funksjoner, sykdommerZygote utvikling
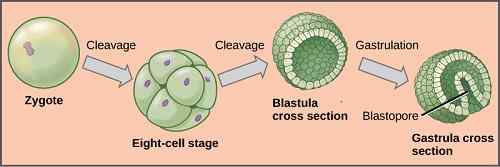
Når zygoten har dannet seg, begynner dette å oppleve en serie endringer og transformasjoner som består av en påfølgende serie mitose som transformerer den til en masse diploide celler kjent som morula.
Utviklingsprosessen som krysser zygoten dekker flere stadier: segmentering, blastulasjon, gastrasjon og organogenese. Hver av dem er av overveiende betydning, siden de spiller en nøkkelrolle i dannelsen av det nye vesenet.
-Segmentering
Dette er en prosess der zygoten opplever mange mitotiske inndelinger, og multipliserer antall celler. Hver av cellene som er dannet fra disse divisjonene er kjent som blastomerer.
Prosessen skjer som følger: Zygoten er delt inn i to celler, igjen er de to delt av å forårsake fire, disse fire i åtte, du er i 16 og til slutt er du i 32.
Den kompakte cellemassen som er dannet, er kjent som morula. Dette navnet skyldes at utseendet ligner på en bjørnebær.
Avhengig av mengden og plasseringen av Vitellus er det fire typer segmentering: holoblastic (total), som kan være like eller ulik; og meroblastisk (delvis), som også kan være like eller ulik.
Holoblastisk eller total segmentering
I denne typen segmentering er hele zygoten segmentert gjennom mitose, noe som resulterer i blastomerer. Nå kan holoblastisk segmentering være av to typer:
- Den samme holoblastiske segmenteringen: I denne typen holoblastisk segmentering er de to første divisjonene langsgående, mens den tredje er ekvatorial. På grunn av dette dannes 8 blastomerer som er like. Disse igjen fortsetter å bli delt gjennom mitose for å danne morulaen. Holoblastisk segmentering er typisk for isolekitos egg.
- Ulik holoblastisk segmentering: Som i all segmentering er de to første divisjonene langsgående, men den tredje er breddegrad. Denne typen segmentering er typisk for mesolecitos egg. I denne forstand dannes blastomerer over hele zygoten, men de er ikke de samme. I zygoten der det er liten mengde vitle, er blastomerer som dannes små og er kjent som mikrometer. Tvert imot, i den delen av zygoten som inneholder rikelig Vitle, kalles blastomerene som stammer fra makromrer.
Meroblastisk eller delvis segmentering
Det er typisk for zygoter som inneholder rikelig vitle. I denne typen segmentering er bare den så -kalt dyrestangen delt. Den vegetative polen er ikke involvert i divisjonen, på en slik måte at en stor mengde vitellus er uten segmentering. På samme måte er denne typen segmentering klassifisert som discoidal og overfladisk.
Meroblastisk diskuidal segmentering
Her opplever bare dyrepolen til zygoten. Resten av dette, som inneholder ganske vitle er ikke segmentert. På samme måte dannes en blastomere -plate som senere vil gi opphav til embryoet. Denne typen segmentering er typisk for telolecitos zygoter, spesielt hos fugler og fisk.
Overfladisk meroblastisk segmentering
I overfladisk meroblastisk segmentering opplever kjernen flere divisjoner, men cytoplasmaet gjør det ikke. På denne måten oppnås flere kjerner, som beveger seg mot overflaten, og distribuerer gjennom hele cytoplasma -dekket. Deretter er cellen grenser som genererer en blastoderm som er perifert, og som omgir vitel som ikke var segmentert, vises. Denne typen segmentering er typisk for leddyr.
-Blastulation
Det er prosessen som følger segmentering. Under denne prosessen blir blastomerer sammen med hverandre og danner veldig nær og kompakte celleforeninger. Gjennom blastulasjon dannes blastula. Dette er en hul struktur, i form av en ball, med et indre hulrom kjent som blastokel.
Struktur av blastula
Blastoderm
Det er det ytre cellelaget som også kalles trofoblast. Det er av vital betydning fordi morkaken og navlestrengen fra det vil bli dannet, viktige strukturer som en utveksling mellom moren og fosteret er etablert.
Det kan tjene deg: Colima Flora og FaunaDet dannes av et stort antall celler som migrerte fra innsiden av morulaen til periferien.
Blastocle
Det er det indre hulrommet til blastocyst. Det dannes når blastomerer migrerer til de ytre delene av morulaen for å danne blastoderm. Sprengningen er okkupert av en væske.
Ebrioblasto
Det er en indre cellemasse, som ligger inne i blastocyst, spesielt i en av dens ender. Fra embryoblasten vil selve embryoet bli dannet. Embryoblasten på sin side består av:
- Hypoblast: Cellelag som er lokalisert i den perifere delen av den primære Vitelino -sekken.
- Epiblast: Cellelag som ligger ved siden av fostervannshulen.
Både epiblast og hypoblast er strukturer av største betydning, siden de så -kallede spirende blader fra dem vil bli utviklet som etter en serie transformasjoner, vil gi opphav til de forskjellige organene som integrerer individet.
Gastulasjon
Dette er en av de viktigste prosessene som oppstår under embryonal utvikling, siden det tillater dannelse av de tre spirminative lagene: endoderm, mesoderm og ektoderm.
Det som skjer under gastruulering er at epiblastceller begynner å spre seg til det er så mange, de må flytte andre steder. På en slik måte at de beveger seg mot hypoblasten, og flytter til og med noen av cellene i dette. Dermed dannes den såkalte primitive linjen.
Umiddelbart oppstår en invaginering, gjennom hvilken cellene i den primitive linjen blir introdusert i retning av blastoklen. På denne måten er et hulrom kjent som arkeistron dannet, som har en åpning, blastoporo.
Slik dannes et bilaminært embryo, bestående av to lag: endoderm og ektoderm. Imidlertid kommer ikke alle levende vesener fra et bilaminært embryo, men det er andre, som mennesket, som kommer fra et trilaminært embryo.
Dette trilaminære embryoet dannes fordi Archentron -cellene begynner å spre seg og til og med plassere mellom ektoderm og endoderm, og gir opphav til et tredje lag, mesoderm.
Endoderm
Fra dette spirende laget dannes epitelet til organene i luftveis- og fordøyelsessystemer, så vel som andre organer som bukspyttkjertel og lever.
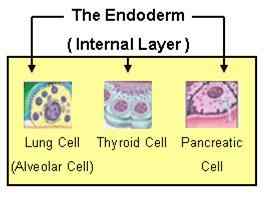 Organer som stammer fra endoderm. Kilde: Endoderm2.PNG: J.Steinbockmaga [Public Domain]
Organer som stammer fra endoderm. Kilde: Endoderm2.PNG: J.Steinbockmaga [Public Domain] Mesoderm
Det gir opphav til bein, brusk og frivillige eller stripete muskler. På samme måte er det dannet organer i sirkulasjonssystemet og andre som nyren, gonadene og myokardiet, blant andre.
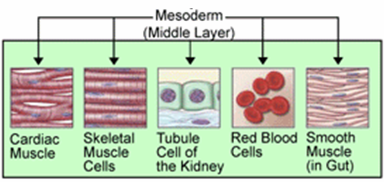 Stoffer avledet fra mesoderm. Kilde: J.Steinbock [Public Domain]
Stoffer avledet fra mesoderm. Kilde: J.Steinbock [Public Domain] Ectoderm
Han er ansvarlig for dannelsen av nervesystemet, huden, neglene, kjertlene (svette og sebaceous), binyremargen og hypofysen.
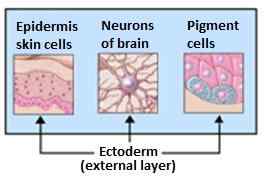 Derivater av ektoderm. Kilde: Ectoderm.PNG: Catmaga [CC BY-SA 3.0 (http: // creativecommons.Org/lisenser/by-SA/3.0/]]
Derivater av ektoderm. Kilde: Ectoderm.PNG: Catmaga [CC BY-SA 3.0 (http: // creativecommons.Org/lisenser/by-SA/3.0/]] Organogenese
Det er prosessen der, fra de spirende lagene og gjennom en serie transformasjoner, hver og en av organene som vil gjøre det nye individet fra sin opprinnelse.
Stort sett er det som skjer her i organogenesen, at stamcellene som er en del av de spirende lagene begynner å uttrykke gener som har funksjonen til å bestemme hvilken type celle som vil opprinnelig.
Avhengig av det evolusjonære nivået i det levende vesenet, vil selvfølgelig organogeneseprosessen være mer eller mindre kompleks.
Referanser
- Carrillo, d., Yaser, l. og Rodríguez, n. (2014). Grunnleggende konsepter om embryonal utvikling i kua. Vaca Reproduksjon: Didaktisk manual for reproduksjon, svangerskap, amming og velvære for den bovine kvinnelige kvinnen. University of Antioquia. 69-96.
- Cruz, r. (1980). Genetiske grunnlag for begynnelsen av menneskelivet. Chilensk pediatri -magasin. 51 (2). 121-124
- López, ca., Garcia, v., Mijares, J., Domínguez, J., Sánchez, f., Álvarez, i. Og Garcia, V. (2013). Gastulasjon: Nøkkelprosess i dannelsen av en ny organisme. Asebir. 18 (1). 29-41
- López, n. (2010). Zygoten av arten vår er en menneskekropp. Person og bioetikk. 14 (2). 120-140.
- Sadler, t. (2001). Langmans medisinsk embryologi. Pan -American Medical Editorial. 8. utgave.
- Ventura, p. Og Santos, m. (2011). Begynnelsen av livet til et nytt menneske fra det vitenskapelige biologiske perspektivet og dets bioetiske implikasjoner. Biologisk forskning. 44 (2). 201-207.
- « Kermés opprinnelse, spill og tradisjoner i Latin -Amerika
- Myose årsaker, patofysiologi og behandlinger »