

Archaea -domenet

- 849
- 241
- Prof. Theodor Gran
 Archaeas illustrasjon
Archaeas illustrasjon Hva er archaea -domene?
Han Archaea -domenet o Archaea Kingdom representerer et av de tre livets domener. Den er sammensatt av veldig like mikroskopiske encellede organismer og samtidig veldig forskjellig fra bakterier og eukaryoter i mange aspekter.
Eksistensen av denne gruppen ble demonstrert nylig, mer eller mindre i slutten av 1970 -buene, også kalt Archeobacteria.
WOEE- og samarbeidspartnere -studier var basert på sammenlignende fylogenetiske analyser mellom sekvensene av ribosomale RNA av alle levende vesener, og forestillingen om de tre domenene opprettholdes, til tross for den enorme mengden sekvenser som fortsetter å bli lagt til kataloger med ribosomal RNA (databaser).
Disse studiene klarte å skimte muligheten for at buene er en brorgruppe av eukaryoter, til tross for deres likheter med prokaryotene (bakterier), slik at de kunne representere den "tapte koblingen" mellom prokaryoter og eukaryoter.
Det antas at disse mikroorganismer representerer mer enn 20% av alle eksisterende prokaryoter i oseaniske farvann, mellom 1 og 5% i jord, og den mest dominerende gruppen i marine sedimenter og geotermiske naturtyper.
Buene er preget av å bo i "ekstreme" forhold som varme kilder, Salinas, miljøer med veldig høye temperaturer og veldig sur pH, ugjestmilde steder der oksygenkonsentrasjon er veldig redusert eller null osv.
Evolusjonær opprinnelse og rFilogenetiske elikasjoner
Det er tre teorier om mulig opprinnelse til de tre livets domener foreslått av Woese og samarbeidspartnere:
- Bakterier divergerte først og dannet en avstamning som produserte buer og eukaryoter
- En "proto-eucariotisk" avstamning divergerte fra en helt probotisk avstamning (den av bakterier og buer)
- Buene divergerte av en avstamning som senere ga opphav til eukaryoter og bakterier
I 1989 foreslo to forskere, Gogarten og Iwabe, uavhengig en måte å fylogenetisk analysere alle levende organismer (en umulig ting å gjøre fra studiene av unike gensekvenser).
Iwabe brukte analysen av sekvensene av gener produsert i "tidlige" hendelser av gentuplikasjon, "forankret" livets tre fra sammenligningen av sekvensen til papegøyegenene som koder for forlengelsesfaktorer.
Forlengelsesfaktorer er GTP -bindende proteiner som deltar i oversettelsen, spesielt i foreningen av overføring RNA -molekyler amioakilert til ribosomer og i translokasjonen av peptidiloverføring.
I henhold til resultatene fra sammenligningene mellom sekvensene til de tre gruppene, er genene som koder for forlengelsesfaktorene i buene mer like de til eukaryote organismer enn de av bakterier.
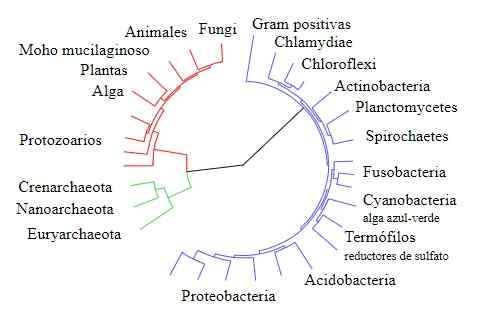 Livets tre
Livets tre Gogarten sammenlignet derimot sekvensene til andre gener produsert ved dupliseringshendelser, spesielt de som koder for type V og type F -underenheter av ATPase -enzymet som finnes i buer/eukaryoter og i bakterier, henholdsvis.
Resultatene oppnådd av Gogarten, som de som er angitt ovenfor, viser at disse genene i buene (som angivelig kom frem fra genetiske dupliseringshendelser) er nærmere relatert til eukaryoter enn med deres bakterielle kolleger.
Disse argumentene ble også støttet av analysen utført av andre forskere mange år senere, som brukte sekvenser fra en annen familie av dupliserte gener, den av aminoacil -synthesaseoverføringen, og styrket unnfangelsen av "nærheten" mellom buene og eukaryoter.
Det kan tjene deg: selenito buljong: hva er, grunnlag, forberedelse, brukDet woese treet
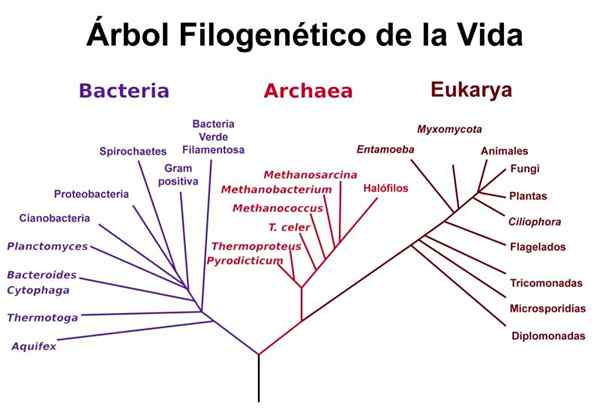 Woese's Life Tree
Woese's Life Tree Woese brukte analysene utført av Gogarten og Iwabe og andre studier relatert til ribosomale RNA -sekvenser for å foreslå sin "versjon" av livets tre, der det er tydelig at buer og eukaryoter utgjør "brødre" -grupper, atskilt fra bakterien selv Selv om ribosomale RNA -sekvenser mellom buer og bakterier er mer like hverandre.
Archaea -domenefunksjonene
Buene er kjent for noen veldig spesielle (egne) egenskaper og også for å presentere "kombinasjoner" av egenskaper som en gang ble antatt var eksklusive for bakterier eller for eukaryote organismer.
- Som bakterier, buer De er prokaryote byråer, Det vil si at inne i det genetiske materialet er ikke låst av en membran (de har ingen kjerne), og det er ingen cytoplasmatiske membranøse organeller.
- De er generelt mikroorganismer som ligner på bakterier, deres DNA er i form av en Sirkulært kromosom og noen mindre sirkulære fragmenter kjent som Plasmider.
- De deler med bakterier tilstedeværelsen av Topoisomerase og turnerer enzymer Lignende, som representerer "indirekte" bevis på "nærheten" når det gjelder den kromosomale strukturen til begge gruppene av organismer.
- Arkaasgener viser imidlertid en stor Homologi med mange eukaryoter, Spesielt med de hvis oppdagelse stammer fra studier med antibiotika.
- Maskineriet til Replikering, transkripsjon og oversettelse av eukaryoter og buer er veldig likt, spesielt med tanke på enzymet ADN -polymerase.
- Dets kodende gener for protein De har ingen introner (Men andre gjør det), i motsetning til eukaryote gener. I tillegg har buer proteiner som ligner histoner assosiert med deres DNA, til stede i eukaryoter og fraværende i bakterier.
- De er preget av tilstedeværelsen av Ether-lipider av isoprenil i cellemembranene, så vel som ved fravær av acylsoler-lipider og en fettsyresynthesase.
- En av underenhetene til RNA -polymerase -enzymet er delt og dets messenger -RNA, så vel som i bakterier, har ikke "capuchas" (fra engelsk kap) På endene 5 '.
- De har et veldig spesifikt utvalg av antibiotikasensitivitet og har Type II Begrensningsenzymer veldig lik de som er beskrevet for bakterier.
- En annen viktig funksjon har å gjøre med det faktum at mye av buene har Cellulær vegg, Men i motsetning til bakterier, er dette ikke sammensatt av peptidoglykan.
Kjennetegn på membranlipider
Arkeiske membranlipider skiller seg betydelig fra de som finnes i bakterier og eukaryote organismer, og dette har blitt ansett som en veldig viktig differensialegenskap.
Hovedforskjellen mellom disse amfipatiske molekylene (med en hydrofil polar ende og en hydrofob apolar) er at foreningen mellom den delen av glyserol og kjeder av fettsyrer i lipidene til buene er gjennom en eterkobling, mens i bakterier og eukaryoter tilsvarer en esterkobling.
En annen viktig forskjell er at buene har lipider med fettsyrer preget av tilstedeværelsen av sterkt forgrenede isoprenile kjeder med metylgrupper, i mellomtiden har eukaryoter og bakterier hovedsakelig fettsyrer av ikke -forgrenede kjeder.
Lipidene til eukaryoter og bakterier er "konstruert" på et glyserolskjelett som fettsyrekjedene er esterifisert i posisjonene som tilsvarer karbonatomer 1 og 2, men i buene inneholder glyseroletene inneholder syrer fett i posisjoner 2 og 3.
Kan tjene deg: Flora og Fauna av Río Negro: Representative arterEn annen forskjell med hensyn til membranlipider har å gjøre med den biosyntetiske ruten, ettersom noen enzymer også er forskjellige i buene.
For eksempel har noen arter av buer et bifunksjonelt overføringsprenilenzym, som er ansvarlig for å tilveiebringe forløpere både for syntese av escuellen og for syntese av glyseril-lipid isaprensoider. I bakterier og eukaryoter blir disse funksjonene utført av separate enzymer.
Klassifisering av buer
I henhold til dataene om sekvensene til de små underenhetene til ribosomalene til buene, har denne gruppen hovedsakelig blitt delt hovedsakelig i to "kanter", som er kjent som kanten Crenarchaeota og kanten Euryarchaeta, hvis medlemmer er fremfor alt dyrket buer In vitro.
Imidlertid har mange av buene som er beskrevet nylig ikke blitt dyrket In vitro Og de er bare fjernt relatert til sekvensene som er blitt isolert fra arten som er opprettholdt i laboratoriene.
Kant Crenarchaeota
 Termofile bakterier, Crenarchaeota Edge
Termofile bakterier, Crenarchaeota Edge Denne gruppen består hovedsakelig av arter av hypertermofile og termacidofile buer, det vil si de kjønnene i buer som bor i ugjestmilde med termiske forhold og ekstrem pH.
Den er sammensatt av en enkelt taksonomisk klasse, som er kjent som Themoprotei, Innenfor er de neste fem taksonomiske ordrene: Acidilobales, Desulfurococcal, Fervidicoccales, Sulfolobales og Termoproteales.
Eksempel på noen av sjangrene som tilhører disse klassene kan være sjangrene Sulfolobus, Desulforococcus, Pyrodictium, Themoproteus og Themofilum.
Kant Euryarchaeta
 Methanosarcina Barkeri Fusaro, Euryarchaota Filo
Methanosarcina Barkeri Fusaro, Euryarchaota Filo Medlemmene av denne gruppen har et litt større økologisk område, og kan finne noen hypertermofil, metenogene, halofile og til og med termofile måler, nimidale buer, svovelreduserende, jernoksidader og noen organrofiler og noen organotrofiler.
Taksonomiske klasser beskrevet for Euriarqueotas er åtte og er kjent som Metanopyri, Methanococci, Methanobacteria, Methanomycrobia, Archaeglobi, Halobacteria, Thermococci og Themoplasmata.
Mange av buene som tilhører denne gruppen er vidt distribuert, og er i jordsmonnene, sedimentene og marine farvannene, så vel som i de ekstreme miljøene som er beskrevet.
Kant Thaumarchaeota
 Axinella -polypoider, Thaumarchaeota Filo. Kilde: Liné1, CC BY-SA 3.0, via Wikimedia Commons
Axinella -polypoider, Thaumarchaeota Filo. Kilde: Liné1, CC BY-SA 3.0, via Wikimedia Commons Denne kanten ble definert relativt nylig, og veldig få av artene som tilhørte den, er blitt dyrket In vitro, Så lite er kjent om disse organismer.
Alle kantmedlemmer får sin energi fra ammoniumoksidasjon og distribueres globalt i ferskvannskropper, jordsmonn, sedimenter og varme kilder.
Philos Korachaeota, Aigachaeota og Garchaeota
 Korachaeota
Korachaeota Noen ekspertforskere på området, basert på analysen av genomiske sekvenser, har nylig bestemt eksistensen av tre ekstra kanter i Archaea -riket, selv om arten som ble foreslått for disse kantene ennå ikke var isolert i et laboratorium.
Generelt er medlemmer som tilhører disse kantene blitt funnet under overflaten av mange land- og marine økosystemer, men også i termisk vann og i dyphavs hydrotermiske systemer.
Ernæring
Flertallet av arkeiske arter med Chimiórophos -organismer, det vil si er i stand til å bruke sterkt reduserte uorganiske forbindelser for å få den energien de trenger for å "flytte" metabolske maskiner, spesielt den som har å gjøre med å puste.
"Spesifisiteten" av de uorganiske molekylene som de bruker som underlag for å produsere energi, avhenger av miljøet der hver enkelt art utvikler.
Andre buer, så vel som planter, alger, briofytter og cyanobakterier, er i stand til å utføre fotosyntese, det vil si at de bruker og konverterer lysenergien til solstråler til nyttig kjemisk energi.
Det kan tjene deg: proteinaminosyrerDet er vist at noen buer lever i magen (vommen) til noen drøvtyggere (blant dem er kyr, sauer, geiter, etc.), så disse beskrives som "gjensidige buer", siden de bruker en del av fiberen som disse dyrene spiser og samarbeider med fordøyelsen av noen av komponentene deres.
Reproduksjon
Akkurat som bakterier er buer encellede organismer hvis reproduksjon utelukkende er aseksuelle. Hovedmekanismene beskrevet fra arten som er opprettholdt In vitro er:
- Binær fisjon, der hver arkea er "del" i to for å stamme to identiske celler
- Geminering eller "fragmentering", der celler gir fra seg "fragmenter" eller "porsjoner" av seg selv som er i stand til å danne nye, genetisk identiske celler.
Habitat
Buene har hovedsakelig relatert til "ekstreme" miljøer, det vil si de naturlige stedene som pålegger alvorlige begrensninger for normal utvikling av levende vesener, spesielt når det gjelder temperatur, pH, saltholdighet, anaerobiose (fravær av oksygen), etc.; Årsaken til at studien din er ekstremt interessant, fordi de har unike tilpasninger.
Imidlertid de nyeste teknikkene for molekylær analyse for identifisering av arter av uaktiverte mikroorganismer (isolert og vedlikeholdt In vitro I et laboratorium) har de tillatt å oppdage tilstedeværelsen av Arch.
Imidlertid er de fleste av buene som er identifisert i naturen klassifisert i henhold til habitatet de okkuperer, og er pårørende i litteraturen begrepene "hypertermofiler", "acidophils" og "ekstreme termacidofiler", "ekstreme halofiler" og "metanogenos".
Miljøene okkupert av hypertermofile buer er de som er preget av veldig høye konstante temperaturer (godt over de "normale" temperaturene som de fleste levende vesener blir utsatt for).
Miljøene der ekstreme acidofiler lever, derimot, er de der pH er veldig lave, og disse kan også skilles ut ved høye temperaturer (ekstreme termacidofiler), i mellomtiden er miljøene til ekstreme halofiler de der konsentrasjonen av salter er veldig høy.
Metanogene buer lever i fravær av oksygen eller anaerobiose, i miljøer der andre molekyler som elektronakseptorer i metabolismen deres kan brukes og er i stand til å produsere metan som et produkt av metabolsk "avfall.".
Eksempler på buerarter
Det er mange kjente arter av buer, men her vil bare noen av dem bli laget.
Ignicoccus Hospitalis og Nanoarchaeum equitans
 Ignicoccus Hospitalis
Ignicoccus Hospitalis Yo. Hospitalis Det tilhører slekten til Crenarqueotas kjent som Ignicoccus og er en kvimiolitotrof organisme som bruker molekylært hydrogen som en elektrondonor for svovelreduksjon. Denne arten har det minste genomet av alle buer beskrevet in vitro så langt.
Yo. Hospitalis Det oppfører seg som en "parasitt" eller "symbiote" av en annen art: Nanoarchaeum equitans. Sistnevnte har ikke blitt dyrket In vitro Og genomet er det minste av alle ubestemte buer som er beskrevet.
Det bor hovedsakelig i marine miljøer og har ikke gener for biosyntese av lipider, aminosyrer, nukleotider eller kofaktorer, så eksperimentelle bevis tyder på at det oppnår disse molekylene takket være dens interaksjon med Yo. Hospitalis.
Acidilobus saccharovorans
Dette er en slags anaerob termacidofil arkea, det vil si at lever i fattige eller helt mangler oksygenmiljøer, med høye temperaturer og pH ekstremt lave. Det ble funnet for første gang i kropper av terrestriske varme kilder i Kamchatka.
Staphylothermus hellenicus
Denne arkeaen tilhører kanten av crenarqueotas, spesielt til rekkefølgen av Desulfurococcal. Det er en hypertermofil heterotrofe arkea (lever i veldig varme miljøer) og krever svovel for å oppnå energi.