

Fosfatidilenolamin
- 2134
- 77
- Jonathan Moe
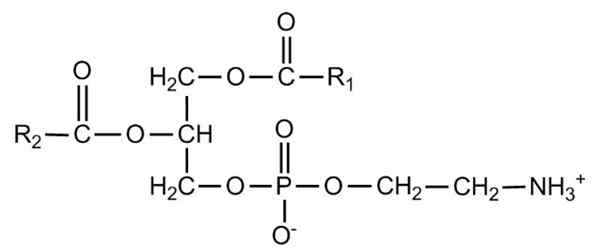
Semi -utviklet formel av fosfatidyletanolamin. Kilde: Zirgouflex, Wikimedia Commons Hva er fosfatidilenolamin?
De fosfatidilenolamin (PE) er en rik glyceofosfolipid i cellemembranene i humant vev. Til tross for dens overflod, avhenger det ikke bare av celletypen, men også av rommet og tiden for den spesifikke cellelivssyklusen som blir vurdert.
Biologiske membraner er barrierer som definerer celleorganismer. Ikke bare har beskyttelses- og isolasjonsfunksjoner, men de er også nøkkelen til etablering av proteiner som krever et hydrofobt miljø for optimal funksjon.
Både eukaryotas og prokaryot.
Glyceofosfolipider er strukturerte amfipatiske molekyler på et L-glyserolskjelett som er esterifisert i SN-1 og SN-2-posisjoner av to fettsyrer i lengde og grad av metningsvariabler. I hydroksylen til SN-3-posisjonen er det forestret seg av en fosfatgruppe, som igjen kan bli med i forskjellige typer molekyler som gir opphav til de forskjellige typer glyserofosfolipider.
I celleverdenen er det et bredt utvalg av glyceofosfolipider, men de mest tallrike er fosfatidylkolin (PC), fosfatidyletanolamin (PE), fosfatidylserin (PS), fosfatidylinitol (Pi), phosfatid (pa), fosfatidylinitol (pi), phosfatid (pa), fosfatidylinitol (pi), pi), pi), pi), fosfatidylinitol (pi), pi), fosfaty (PS), fosfatidylinitol (Pi), fosfatid (PS), fosfatidylinitol (Pi), fosfatid (PS), fosfatidylinitol (Pi), fosfatol), ps), fosfatidylinitol (Pi), fosfat, (CL).
Struktur
Strukturen til fosfatidilenolamin ble oppdaget av Baer og samarbeidspartnere i 1952. Ettersom det er blitt bestemt eksperimentelt for alle glyserofosfolipider, dannes fosfatidyletanolamin av et glyserolmolekyl sterifisert i SN-1 og SN-2-posisjoner med fettsyrekjeder mellom 16 og 20 karbonatomer.
De sterifiserte fettsyrene i SN-1-hydroksylen er generelt mettede (uten dobbeltbindinger) med maksimal lengder på 18 karbonatomer, mens de forente kjedene i SN-2-stillingen er av større lengde og med en eller flere umettethet (dobbeltkoblinger).
Det kan tjene deg: soppsirkulasjon: næringsstoffer, stoffer, osmoreguleringMetningsgraden av disse kjedene bidrar til membranens elastisitet, som har stor innflytelse på innsetting og kidnapping av protein i Bilay.
Fosfatidyletanolamin regnes som en ikke -lamellær glyceofosfolipid, siden den har en konisk geometrisk form. Denne formen er gitt av den lille størrelsen på din polar- eller "hode" -gruppe, i forhold til den for fettsyrekjeder som omfatter de hydrofobe "køene".
"Hodet" eller den polare gruppen av fosfatidyletanolamin er zwitterionisk karakter, det vil si at den har grupper som kan være positive og negativt ladet under visse pH -forhold.
Denne funksjonen lar deg etablere hydrogenbindinger med en stor mengde aminosyreavfall, og dens belastningsfordeling er en essensiell determinant for topologien til domenene til mange omfattende membranproteiner.
Biosyntese
I eukaryote celler er syntesen av strukturelle lipider geografisk begrenset, og er hovedstedet for biosyntese.
Det er fire uavhengige biosyntetiske ruter for produksjon av fosfatidyletanolamin: (1) CDP-stanolaminruten, også kjent som Kennedy-ruten. (2) PSD -ruten for fosfatidylserin -dekarboksylering (PS). (3) acilatasjonen av glatt.
Kennedy Route
Biosyntesen av fosfatidyletanolamin av denne ruten er begrenset til ER, og det er vist at i hamsterleverceller er det hovedveien for produksjon. Den består av tre påfølgende enzymatiske trinn katalysert av tre forskjellige enzymer.
Kan tjene deg: Flora og Fauna of Campeche: Representative arterI det første trinnet er det fosfoetanolamin og ADP takket være virkningen av kinase etanoolamin, som katalyserer den ATP -avhengige fosforylering av etanoolamin.
I motsetning til planter, er verken pattedyr eller gjær i stand til å produsere dette underlaget, så det må konsumeres i kostholdet eller oppnås fra nedbrytning av fosfatidyletalamin eller sfærisk fosfatidile eller sphygosinmolekyler.
Fosfoetanolamin brukes av CTP: fosfoetanolamin citidiltransferase (ET) for å danne den høye energiforbindelsen CDP: etanoolamin og en uorganisk fosfat.
1,2-diacylglycerol etanoolamin fosfotransferase (ETP) bruker energien som er inneholdt i CDP-enanolaminlenken for å kovalent forene etanolamin til et diacylglycerolmolekyl.
PSD -rute
Denne ruten fungerer i både prokaryoter og gjær og pattedyr. I bakterier forekommer det i plasmamembranen, men i eukaryoter foregår det i et område av det endoplasmatiske retikulumet som har et nært forhold til mitokondriell membran.
Hos pattedyr katalyseres ruten av et enkelt enzym, fosfatidylserin disarboxylase (PSD1p), som er innebygd i mitokondriell membran, hvis gen er kodet av kjernen. Reaksjonen innebærer dekarboksylering av PS til fosfatidyletanolamin.
De resterende to rutene (acilatisering av glatt-PE og kalsiumavhengig utveksling av polargruppe) forekommer i endoplasmatisk retikulum, men bidrar ikke vesentlig til den totale produksjonen av fosfatidilenolamin i eukaryote celler i eukaryote celler.
Funksjoner
- Glyceophosphalipids har tre hovedfunksjoner i cellen, hvorav strukturell, energilagring og celleskiltfunksjoner er.
- Fosfatidilenolamin er assosiert med forankring, stabilisering og folding av flere membranproteiner, så vel som de nødvendige konformasjonsendringene for drift av mange enzymer.
- Det er eksperimentelle bevis som foreslår fosfatidyletanolamin som en avgjørende glyceofosfolipid i det sene stadiet av telofasen, under dannelsen av den kontraktile ringen og etableringen av ramoplasten som tillater deling av membranen til de to cellene.
- Den har også en viktig funksjon i alle fusjons- og fisjonsprosesser (union og separasjon) av membranene til både endoplasmatisk retikulum og Golgi -apparatet. I OG. coli Det er bevist at fosfatidyletanolamin er nødvendig for riktig folding og funksjon av permease laktoseenzymet, så det har blitt antydet at det har en rolle som molekylær "chaperone".
- Fosfatidilenolamin er hoveddonoren til etanolaminmolekylet som er nødvendig for den post-translasjonelle modifiseringen av mange proteiner, så som GPI-forankringer.
- Denne glyceofosfolipiden er forløperen for mange molekyler med enzymatisk aktivitet. I tillegg kan molekyler avledet fra metabolismen, så vel som diCilglycerol, fosfatidsyre og noen fettsyrer, fungere som andre budbringere. I tillegg er det et viktig underlag for fosfatidylkolinproduksjon.
Referanser
- Gibellini, f., & Smith, t. K. Kennedy Pathway-of Novo-syntese av fosfathidyletanolamin og fosfatidylkolin. IUBMB LIV.
- Harayama, t., & Riezman, h. Forstå mangfoldet av membranlipidsammensetning. Naturen vurderer molekylær cellebiologi.