

Lipogeneseegenskaper, funksjoner og reaksjoner
- 1431
- 250
- Marius Aasen
De lipogenese Det er den viktigste metabolske ruten som langkjedede fettsyrer blir syntetisert fra overflødig karbohydrater i kostholdet. Disse fettsyrene kan inkorporeres i triglyserider ved hjelp av sterifisering til glyserolmolekyler.
Under normale forhold oppstår lipogenese i lever- og fettvevet og regnes som en av de viktigste skattebetalerne for å opprettholde triglyseridhomeostase i blodserum.
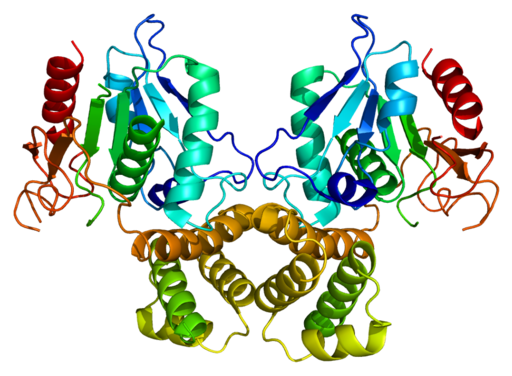
 Human Syntasa Fat Structure (FASN) (Kilde: EMW
Human Syntasa Fat Structure (FASN) (Kilde: EMW[CC By-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] Via
Wikimedia Commons)
Triglyserider er det viktigste energireservoaret i kroppen, og energien i disse blir trukket ut takket være en prosess kjent som lipolyse som i motsetning til lipogenese består av separasjon og frigjøring av glyserolmolekyler og fettsyrer mot blodstrømmen.
Den frigjorte glyserol fungerer som et underlag for den glukoneogene ruten, og fettsyrer kan transporteres til andre gratis rom med serumalbumin.
Disse fettsyrene blir fanget opp av nesten alle vev med unntak av hjernen og erytrocytter, deretter blir de sterifisert til triacylglyserol igjen for å oksideres som drivstoff eller lagres som en energireserve.
Fett -rike dietter er de viktigste årsakene til overvekt, siden kalorioverskuddet må lagres og fettvev må utvide seg for å imøtekomme både overflødige lipider og de som er syntetisert endogent.
[TOC]
Egenskaper og funksjoner
I menneskekroppen, for eksempel, oppstår fettsyrer en av biosyntetiske prosesser fra acetyl-CoA eller som et produkt av den hydrolytiske prosessen av membranfett og fosfolipider.
Mange pattedyr klarer ikke å syntetisere noen fettsyrer, noe som gjør disse essensielle komponentene i kostholdet.
Hovedfunksjonen til lipogenese har å gjøre med lagring av energi i form av fett (lipider) som oppstår når du konsumerer en større mengde karbohydrater enn de som kroppen trenger, til og med overskrider leverens lagringskapasitet i glykogen.
Kan tjene deg: gastrine: egenskaper, struktur, produksjon, funksjonerLipider som er syntetisert med denne ruten, lagres i det hvite fettvevet, hovedstedet for lagring av lipider i kroppen.
I alle kroppsceller forekommer lipogenese, men fettvev og leveren er de viktigste syntese stedene. Denne ruten er gitt i cellens cytoplasma, mens oksidasjon av fettsyrer forekommer i mitokondrielle rom.
Lipogenese og påfølgende syntese av triglyserider blir fulgt av syntese og sekresjon av lipoproteiske partikler med svært lav tetthet kjent som VLDL -partikler (engelsk Varier lipoprotein med lav tetthet), som er i stand til å komme inn i blodomløpet.
Både VLDL -partikler og triglyserid.
Reaksjoner
Strømmen av karbonatomer fra glukosen som er til stede i karbohydrater til fettsyrer moduleres av lipogenese og inkluderer en serie perfekt koordinerte enzymatiske reaksjoner.
1-Glykolytisk rute i cytosolen til cellene er ansvarlig for å behandle glukosen som kommer inn fra blodomløpet for å produsere pyruvat, som omdannes til acetyl-CoA, i stand til å gå inn i Krebs-syklusen i mitokondriene, hvor sitratet oppstår.
2-Det første trinnet på den lipogene ruten består i konvertering av sitrat som forlater mitokondriene i acetyl-CoA ved virkningen av et enzym kjent som ATP-sitrat Liasa (Acyy).
3-Det resulterende acetyl-CoA er karboksylert for å danne malonyl-CoA, en acetyl-CoA karboksylase (Aqueca).
4-Den tredje reaksjonen er reaksjonen som pålegger det begrensende trinnet i hele ruten, det vil si den tregeste reaksjonen, og består i konvertering av malonyl-CoA til palmitat av en enzymfettsyresyntase (FAS) (FAS).
Det kan tjene deg: Half Cary Blair: Foundation, Preparation and Uses5-andre nedstrøms reaksjoner hjelper til med å konvertere palmitatet til andre mer komplekse fettsyrer, men palmitatet er imidlertid hovedproduktet av lipogenese av novo.
Fettsyresyntese
Syntesen av fettsyrer hos pattedyr begynner med den komplekse fettsyresyntasen (FAS), et multifunksjonelt og multi -knusingskompleks i cytosol som syntetiserer palmitatet (et 16 karbonmettet fettsyre). For denne reaksjonen bruker han, som nevnt allerede, malonyl-CoA som en karbon-giver og NADPH som en kofaktor.
Underenhetene til homodimeren av Fas katalyserer syntesen og forlengelsen av fettsyrer to karbonatomer samtidig. Disse underenhetene har seks forskjellige enzymatiske aktiviteter: Acetyltransferase, B-Cetoacil Syntasa, Malonyl Transferas.
Ulike medlemmer av en familie av forlengelsesprotein av veldig langkjede fettsyrer (ELOVL) er ansvarlige for å forlenge fettsyrene produsert av FAS. Nedstrøms er andre enzymer som er ansvarlige for innføring av dobbeltbindinger (desaturering) i kjeder av fettsyrer.
Regulering
Tallrike patofysiologiske forhold har å gjøre med den mangelfulle reguleringen av den lipogene ruten, siden uregelmessigheter i samme avbrytelseslegeme lipid homeostase.
Et kosthold rik på karbohydrater aktiverer leverlipogenese, men det har vist seg at det ikke bare er mengden karbohydrater som er inntatt, men også typen karbohydrater.
Eksperimentelle data viser for eksempel at enkle sukker som fruktose har mye kraftigere effekter på aktivering av lever lipogenese enn andre mer komplekse karbohydrater.
Glukolitisk glukosemetabolisme representerer en stor kilde til karbon for fettsyresyntese.
Glukose induserer også ekspresjonen av enzymene som er involvert i den lipogene ruten gjennom unionsproteiner til karbohydratresponselementer.
Kan tjene deg: San Luis Potosí Flora og Fauna: Mer representative arterBlodsukkernivået stimulerer også ekspresjonen av disse enzymene ved stimulering av insulinfrigjøring og hemming av glukagon -frigjøring i bukspyttkjertelen. Denne effekten styres via proteinet av binding til det regulatoriske elementet i sterol 1 (SREBP-1) i leverceller og adipocytter.
Andre reguleringsveier har mye å gjøre med det endokrine systemet og forskjellige hormoner indirekte relatert til uttrykk for mange av de lipogene enzymer.
Referanser
- Ameer, f., Scandiuzzi, l., Hasnain, s., Kalbacher, h., & Zaidi, n. (2014). Av novo lipogenese i helse og sykdom. Metabolisme, 0-7.
- Lodhi, i. J., Wei, x., & Semenkovich, C. F. (2011). Lipoexpedy: Fra Novo Lipogenesis som en metabolsk signalsender. Trender i endokrinologi og metabolisme, 22(1), 1-8.
- Mathews, c., Van Holde, K., & Ahern, K. (2000). Biokjemi (3. utg.). San Francisco, California: Pearson.
- Nelson, d. L., & Cox, m. M. (2009). Lehninger prinsipper for biokjemi. Omega -utgaver (5. utg.).
- Samuel, v. T. (2011). Fruktose indusert lipogenese: fra sukker til fett til insulinresistens. Trender i endokrinologi og metabolisme, 22(2), 60-65.
- Scherer, t., Hare, J. ENTEN., Diggs-Andrews, k., Schweiger, m., Cheng, f., Lindtner, ca.,... Buettner, C. (2011). Hjerneinsulin kontrollerer fettvev lipolyse og lipogenese. Cellemetabolisme, 1. 3(2), 183-194.
- Schutz, og. (2004). Kostholdsfett, lipogenese og energibalanse. Fysiologi og atferd, 83, 557-564.
- Stabil, m. S., & Ntambi, j. M. (2010). Genetisk kontroll av novo lipogenese: rolle i kostholdsindusert overvekt. Kritiske gjennomganger i biokjemi og molekylærbiologi, Fire fem(3), 199-214.
- Zaidi, n., Lupien, l., Kuemmerle, n. B., Kinlaw, w. B., Swinnen, J. V, & Smans, K. (2013). Lipogenese og lipolyse: Stiene utnyttet av kreftcellene for å skaffe fettsyrer fettsyrer. Fremgang i lipidforskning, 52(4), 585-589.
- « Aktive læringsegenskaper, stiler og strategier
- Moderne filosofiegenskaper, stadier og representanter »