

Nukleosom
- 1058
- 22
- Prof. Joakim Johansen
Nukleosom er den grunnleggende enheten for organisering av DNA i eukaryote celler Hva er nukleosom?
Han Nukleosom Det er den grunnleggende DNA -emballasjeenheten i eukaryote organismer. Derfor utgjør det det minste kromatinkompresjonselementet.
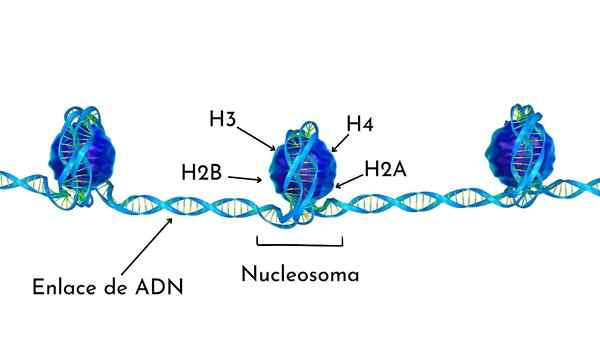
Nukleosomet er bygget som en proteinoktamer som kalles histoner, eller trommelformet struktur som omtrent 140 NT DNA rulles, og gir nesten to komplette runder, som det kan sees på bildet.
Det anses at noen ekstra DNA -NT -er er en del av nukleosomet, og er DNA -fraksjonen som tillater fysisk kontinuitet mellom en nukleosom og en annen i mer komplekse kromatinstrukturer (for eksempel kromatinfiber på 30 nm).
Histonkoden var en av de første bedre epigenetiske kontrollelementene.
Nukleosomfunksjoner
Nukleosomer tillater:
- DNA -emballasje for å imøtekomme den begrensede kjerneplassen.
- De bestemmer partisjonen mellom kromatinet som er uttrykt (eukromatin) og det stille kromatinet (heterokromatin).
- Organisere alt kromatin både romlig og funksjonelt i kjernen.
- De representerer underlaget til de kovalente modifikasjonene som bestemmer ekspresjonen og ekspresjonsnivået, av genene de koder for proteiner gjennom den så -kallede histonkoden.
Sammensetning og struktur
I sin mest grunnleggende forstand er nukleosomer sammensatt av DNA og proteiner. DNA kan være, praktisk talt ethvert dobbeltbånd -DNA som er til stede i kjernen i den eukaryote cellen, mens alle nukleosomale proteiner tilhører settet med proteiner kalt histoner.
Kan tjene deg: celledifferensieringHistonene er små proteiner og med en høy belastning med basisk aminosyreavfall, som gjør det mulig å motvirke den høye negative belastningen av DNA og etablere en effektiv fysisk interaksjon mellom de to molekylene uten å nå stivheten til den kovalente kjemiske bindingen.
Histonene danner en trommel -veioktameter med to eksemplarer eller monomerer av hver av H2A, H2B, H3 og H4 histoner.
DNAet gir nesten to komplette svinger på sidene av oktameren og fortsetter deretter med en brøkdel av Linker -DNA som er assosiert med histonen H1, for å gå tilbake til to komplette svinger i en annen Histona Octa.
Oktametersettet, tilknyttet DNA, og dets tilsvarende linker -DNA, er et nukleosom.
 Deler av et nukleosom
Deler av et nukleosom Kromatinkomprimering
Genomisk DNA utgjøres av ekstremt lange molekyler (mer enn en meter i tilfelle av mennesket, med tanke på alle dets kromosomer), som må være bestemmende og organisert i en ekstremt liten kjerne.
Det første trinnet i denne komprimeringen utføres gjennom dannelse av nukleosomer. Bare med dette trinnet er DNA komprimert omtrent 75 ganger.
Dette gir opphav til en lineær fiber som de påfølgende nivåene av kromatinkomprimering er bygget: 30 nm fiber, båndene og båndene til båndene.
Når en celle er delt, enten ved mitose eller med meiose, er den siste komprimeringsgraden henholdsvis mitotisk eller meiotisk kromosom.
Histonkoden og genetisk uttrykk
Det faktum at histonoktamer og DNA interagerer elektrostatisk elektrostatisk, uten å miste fluiditeten som kreves for å gjøre nukleosomer dynamiske elementer av komprimering og nedbrytning av kromatin.
Kan tjene deg: cellelysisMen det er et enda mer overraskende interaksjonselement: endene n terminalene til histonene blir utsatt utenfor interiøret i oktameteren, mer kompakt og inert.
Disse ytterpunktene samhandler ikke bare fysisk med DNA, men lider også en serie kovalente modifikasjoner som graden av komprimering av kromatin og uttrykk for det tilhørende DNA vil avhenge.
Settet med kovalente modifikasjoner, når det gjelder type og antall, blant annet, er samlet kjent som histonkoden.
Disse modifikasjonene inkluderer fosforylering, metylering, acetylering, ubiquitinering og høyt.
Hver endring vil bestemme uttrykket eller ikke for det tilhørende DNA, samt graden av komprimering av kromatin, i forbindelse med andre endringer i samme molekyl eller i avfall fra andre histoner, spesielt av H3 H3.
Som en generell regel har det blitt sett for eksempel at hypermetilert og hypacetlated histoner bestemmer at tilknyttet DNA ikke er uttrykt og at kromatin presenteres i en mer kompakt tilstand (heterokromatisk, og følgelig inaktiv).
Tvert imot er eukromatisk DNA (mindre kompakt og genetisk aktivt) assosiert med kromatin hvis histoner er hypercetilated og hypometylisert.
Eukromatin og heterokromatin
Histonas 'kovalente modifikasjonsstatus kan bestemme graden av uttrykk og komprimering av lokalt kromatin.
På globale nivåer er kromatinkomprimering like regulert av de kovalente modifikasjonene av histoner i nukleosomer.
Det er for eksempel påvist at konstitutiv heterokromatin (som aldri kommer til uttrykk, og er tettpakket), har en tendens til å være plassert til kjernearket, og etterlater kjernefysiske porer gratis.
Kan tjene deg: kalsiumpumpe: funksjoner, typer, struktur og driftFor sin del gjør konstitutiv eukromatin (som alltid kommer til uttrykk, slik som det som inkluderer cellevedlikeholdsgener, og er lokalisert i slappe kromatinregioner), i store bånd som utsetter DNA for å bli transkribert til transkripsjonsmaskineriet.
Andre regioner med genomisk DNA svinger mellom disse to tilstandene, avhengig av utviklingstid for organismen, av vekstbetingelser, celleidentitet, etc.
Andre funksjoner
For å oppfylle sin cellulære utvikling, uttrykk og vedlikeholdsplan, må genomene til eukaryote organismer finnelig regulere når og hvordan deres genetiske potensialer skal manifestere.
Med utgangspunkt i informasjonen som er lagret i genene sine, er disse lokalisert i kjernen i private regioner som bestemmer deres transkripsjonsstatus.
Derfor kan vi si at en annen av de grunnleggende artiklene av nukleosomer er organisasjonen eller arkitekturen til kjernen som huser dem.
Denne arkitekturen er arvet og fylogenetisk bevart takket være eksistensen av disse modulære elementene i informasjonsemballasje.
Referanser
- Brooker, r. J. Genetikk: Analyse og prinsipper. McGraw-Hill Higher Education.
- Goodenough, u. W. Genetikk. W. B. Saunders co. Ltd.