

Polyploidy -typer, hos dyr, hos mennesker, i planter
- 2384
- 254
- Markus Fredriksen
De Polyploidi Det er en type genetisk mutasjon som består i tillegg av en komplett begavelse (komplette spill) av kromosomer til cellekjernen, som utgjør homologe jevnaldrende. Denne typen kromosomal mutasjon er den vanligste av euploidias og er preget av organismen har tre eller mer komplette kromosomspill.
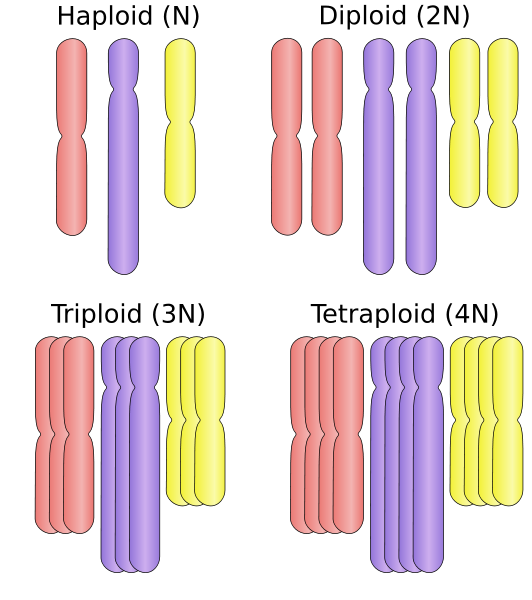
En organisme (diploid normalt = 2n) regnes som polyploid når den skaffer seg ett eller flere komplette sett med kromosomer. I motsetning til spesifikke mutasjoner, kromosomale investeringer og duplikasjoner, er denne prosessen i stor skala, det vil si at den skjer på komplette kromosomer.
Kilde: haploid_vs_diploid.SVG: Ehambergderive Work: Ehamberg [CC By-SA 3.0 (https: // creativecommons.Org/lisenser/by-SA/3.0)] I stedet for å være haploid (n) eller diploid (2n), kan en polyploid organisme være tetraploid (4n), oktoploid (8n) eller høyere. Denne mutasjonsprosessen er ganske hyppig i planter og er rart hos dyr. Denne mekanismen kan øke genetisk variabilitet i de sessile organismer som ikke er i stand til å bevege seg fra miljøet.
Polyploidi er av stor betydning i evolusjonsmessige termer i visse biologiske grupper, der det utgjør en hyppig mekanisme for å generere nye arter siden kromosombelastning er en arvelig tilstand.
[TOC]
Når polyploidien oppstår?
Forstyrrelsene i antall kromosomer kan forekomme både i naturen og i populasjoner som er etablert i laboratoriet. De kan også induseres med mutagent middel som matter. Til tross for den utrolige presisjonen av meiose, oppstår kromosomavvik og er mer vanlig enn man kanskje tror.
Polyploidi oppstår som et resultat av noen endringer som kan oppstå under meiose, enten i den første meiotiske divisjonen eller under profasen, der de homologe kromosomene er organisert i par for å danne tetrader og en ikke-disyunksjon av sistnevnte skjer under sistnevnte under Anafase i.
Utseende av nye arter
Polyploidi er viktig siden det er et utgangspunkt for å forårsake nye arter. Dette fenomenet er en viktig kilde til genetisk variasjon, siden det gir hundrevis eller tusenvis av dupliserte loci som vil være fritt til å oppnå nye funksjoner.
I planter er det spesielt viktig og ganske utbredt. Det anslås at mer enn 50% av blomsterplantene har sin opprinnelse fra polyploidi.
I de fleste tilfeller skiller polyploider seg fysiologisk fra originale arter, og på grunn av dette kan de kolonisere miljøer med nye egenskaper. Mange viktige arter i landbruket (inkludert hvete), er polyploider av hybrid opprinnelse.
Typer polyploidi
Polyloidia kan klassifiseres i henhold til antall komplette sett eller kromosomale spill som er til stede i cellekjernen.
I denne forstand er en organisme som inneholder "tre" kromosomer "triploid", "tetraploid" hvis den inneholder 4 kromosomer, pentaploid (5 spill), heksaploidae (6 spill), heptoploid (syv kamper), octoploid (åtte kamper),, heptoploid (syv spill), oktoploid (åtte spill),,, heptoploid (syv spill), octoploid (åtte spill), Nonaploidae (ni kamper), avskrekke (10 kamper), og så videre.
Kan tjene deg: Økologisk isolasjon: Hva er, mekanisme og eksemplerPå den annen side kan polyploidier også klassifiseres i henhold til opprinnelsen til kromosomale begavelser. I denne ideens rekkefølge kan en organisme være: autopoliploid eller alopoliploid.
En autopoliploid inneholder flere spill av homologe kromosomer avledet fra samme individ eller et individ som tilhører samme art. I dette tilfellet dannes polyploider av foreningen av ikke -reduserte gameter av genetisk kompatible organismer som er klassifisert som den samme arten.
En allopoliploid er at organisme som inneholder spill av ikke -homologe kromosomer på grunn av hybridisering mellom forskjellige arter. I dette tilfellet oppstår polyploidi etter hybridisering mellom to beslektede arter.
Polyploidi hos dyr
Hos dyr er polyploidi sjelden eller sjelden. Den mest utbredte hypotesen som forklarer den lille frekvensen av polyploide arter hos høyere dyr, er at dens komplekse mekanismer for seksuell bestemmelse avhenger av en veldig delikat balanse i antall kjønnskromosomer og autosomer.
Denne ideen har blitt opprettholdt til tross for de akkumulerte bevisene til dyr som eksisterer som polyploid. Det observeres generelt i lavere dyregrupper som ormer og et bredt utvalg av plalmintos, der individer vanligvis har både mannlige og kvinnelige gonader som letter selvfertilisering.
Arten som presenterer denne siste tilstanden kalles autokompatible hermafroditter. På den annen side kan det også forekomme i andre grupper hvis kvinner kan gi avkom uten befruktning, gjennom en prosess som kalles parthenogenese (som ikke innebærer en normal meiotisk seksuell syklus)
Under parthenogenese produseres etterkommere i utgangspunktet ved mitotisk inndeling av foreldreceller. Her er mange arter av virvelløse dyr som biller, isopoder, møll, reker, flere grupper av arachnider og noen arter av fisk, amfibier og krypdyr inkludert.
I motsetning til planter, utgjør spesifikasjonen gjennom polyploidi hos dyr en eksepsjonell hendelse.
Eksempler hos dyr
Gnageren Tympanoctomys Barrerae Det er en tetraploid art som har 102 kromosomer per somatisk celle. Den presenterer også en "gigas" -effekt på sædcellene. Denne alopoliploide arten stammet sannsynligvis fra forekomsten av flere hybridiseringshendelser av andre gnagerarter som for eksempel som Octomys Mimax og Pipanacoctomys aureus.
Polyploidi hos mennesker
Polyploidi er sjelden i virveldyr og anses som irrelevant i diversifisering av grupper som pattedyr (i motsetning til planter) på grunn av forstyrrelsene som oppstår i kjønnsbestemmelsessystemet og i dosekompensasjonsmekanismen.
Det anslås at fem av hver 1000 mennesker blir født med alvorlige genetiske defekter som kan tilskrives kromosomale anomalier. Et enda større antall embryoer med kromosomfeil aborteres spontant og mange når aldri fødselen.
Kan tjene deg: Transposoner: Hva er, typer, egenskaperHos mennesker anses kromosomale polyploider som dødelige. I somatiske celler som hepatocytter er imidlertid omtrent 50% av disse normalt polyploid (tetraploid eller oktaploid).
De mest oppdagede polyploidiene i våre arter er fulle triploidier og tetraploidier, i tillegg mixloid diploid/triploid (2N/3N) og diploid/tetraploid (2N/4N).
Hos sistnevnte sameksisterer en populasjon av normale diploide celler (2N), med en annen som har 3 eller flere haploide multipler av kromosomer, for eksempel: triploid (3n) eller tetraploid (4N).
Triploidies og tetraplodies hos mennesker er ikke levedyktige på lang sikt. I de fleste tilfeller er det rapportert om død i de fleste tilfeller eller til og med noen dager etter fødselen fra mindre enn en måned til maksimalt 26 måneder.
Polyploidi i planter
Eksistensen av mer enn ett genom i den samme kjernen har spilt en viktig rolle i plantenes opprinnelse og utvikling, kanskje den viktigste cytogenetiske endringen i plantespesiasjon og evolusjon. Plantene var inngangsdøren til kunnskapen om celler med mer enn to kromosomer per celle per celle.
Fra begynnelsen av kromosomtallene ble det observert at et bredt utvalg av ville og dyrkede planter (inkludert noen av de viktigste) er polyploide. Nesten halvparten av arten av angiospermer (planter med blomster) er polyploider, på samme måte de fleste bregner (95%) og et bredt utvalg av moser.
Tilstedeværelsen av polyploidi i gymnospermsplanter er sjelden og svært varierende i angiospermsgrupper. Generelt sett har det blitt påpekt at polyploide planter er svært tilpasningsdyktige, og kan okkupere naturtyper som deres diploide forfedre ikke kunne. I tillegg akkumulerer polyploide planter med mer genomiske kopier større "variabilitet".
Innen planter hadde kanskje alopoliploider (mer vanlig i naturen) en grunnleggende rolle i spesiasjons- og adaptiv stråling av mange grupper.
Forbedring av hagebruk
I grønnsaker kan polyploidi stamme fra flere forskjellige fenomener, og være kanskje de hyppigste feilene under meioseprosessen som gir opphav til diploide gameter.
Mer enn 40% av de dyrkede plantene er polyploide, inkludert alfalfa, bomull, potet, kaffe, jordbær, hvete blant andre, uten at det er et forhold mellom domestisering og polyploidi av planter.
Siden madrass ble implementert som et middel for å indusere polyploidi, ble den brukt i avlingsplanter i utgangspunktet av tre grunner:
-For å generere polyploidi i visse arter av betydning, for eksempel et forsøk på å få bedre planter, fordi det i polyploider vanligvis er en fenotype der det er en bemerkelsesverdig vekst av størrelse "gigas" fordi det er et større antall celler. Dette har gjort det mulig.
Kan tjene deg: genstrømning-For polyploidisering av hybrider og at de gjenvinner fruktbarhet slik at den blir redesignet eller syntetisert av noen arter.
-Og til slutt som en måte å overføre gener mellom arter med forskjellig grad av ploidi eller innenfor samme art.
Eksempler i planter
Innen planter er en naturlig polyploid av stor betydning og spesielt interessant hvete av brød, Triticum aestibum (Hexaploid). Sammen med rug, en polyploid kalt "triticale", en allpoliploid med den høye produktiviteten til hvete og robusthet av rug, som har stort potensiale.
Hvete i dyrkede planter har vært bemerkelsesverdig essensielt. Det er 14 arter av hvete som har utviklet seg ved alopolyloidi, og danner tre grupper, en av 14, en annen av 28 og en siste av 42 kromosomer. Den første gruppen inkluderer den eldste arten av slekten T. Monococcum og T. Boooticum.
I den andre gruppen består av 7 arter og stammer tilsynelatende fra hybridiseringen av T. Boooticum Med en slags vill koke fra en annen sjanger som heter Aegilops. Kryssingen produserer en steril kraftig hybrid som ved duplisering av kromosomer kan resultere i en fruktbar allotraploid.
Den tredje gruppen på 42 kromosomer er der brødets hvete er, som sannsynligvis har sin opprinnelse gjennom hybridisering av en stubbart med en annen art av arten av Aegilops etterfulgt av en kromosomisk komplement duplisering.
Referanser
- Alcántar, J. P. (2014). Polyploidi og dens evolusjonære betydning. Utsteder mangel og teknologi, 18: 17-29.
- Ballesta, f. J. (2017). Noen bioetiske hensyn i forhold til eksistensen av tilfeller av mennesker med fullstendig tetraploidi eller triploidy. Studia Bioethica, 10(10): 67-75.
- Castro, s., & Loureiro, J. (2014). Reproduksjonens rolle ved opprinnelsen og utviklingen av polyploide planter. Ecosystems Magazine, 23(3), 67-77.
- Freeman, S og Herron, J. C. (2002). Evolusjonsanalyse. Pearson Education.
- Hichins, c. F. Yo. (2010). Genetisk og geografisk opprinnelse til den tetraploide gnagere tympanoctomys barrerae (Octodontidae), basert på analysen av mitokondrielle cytochondriale sekvenser (Avhandling doktorgrad, Institute of Ecology).
- Hickman, ca. P, Roberts, L. S., Keen, s. L., Larson, a., I'anson, h. & Eisenhour, d. J. (2008). Integrerte priorms av zoologi. New York: McGraw-Hill. 14th Utgave.
- Pimentel Benítez, h., Lantigua Curz, til., & Quiñones maza, eller. (1999). Diploid-tetraploid mixoploidy: Første rapport i vårt miljø. Cuban Pediatrics Magazine, 71(3), 168-173.
- Schifino-Wittmann, m. T. (2004). Polyploidia e SEU Impact Na Origin og Evolução Das Wild and Cultivated Plants. Brasiliansk magasin for agrociencia, 10(2): 151-157.
- Suzuki, d. T.; Griffiths, a. J. F.; Miller, J. H & Lewontin, R. C. (1992). Introduksjon til genetisk analyse. McGraw-Hill Inter-American. 4th Utgave.
- « Euploidía opprinnelse, typer og konsekvenser
- Chamelaucium uncinatum -egenskaper, habitat, omsorg »