

Tilacoid

- 2982
- 243
- Anders Mathisen
Tilacoid i en kloroplast Hva er tilacoider?
De Tilacoid De er rom i form av flate poser som ligger i kloroplaster i planteceller, i cyanobakterier og alger. De er vanligvis organisert i en struktur som kalles grana -plural Granum- og ser ut som en haug med mynter.
Tilacoider regnes som det tredje kloroplastmembransystemet, bortsett fra den indre og ytre membranen til nevnte organelle. Membranen til denne strukturen skiller seg inne i tilacoid av kloroplaststroma, og har en serie pigmenter og proteiner involvert i metabolske veier.
I tilacoider er det essensielle biokjemiske reaksjoner for fotosyntese, prosess der planter tar sollys og forvandler det til karbohydrater. Spesielt har de det nødvendige maskineriet forankret til membranen for å utføre sollysavhengig fase, der lyset er fanget og omdannet til energi (ATP) og NADPH.
Kjennetegn på tilacoider
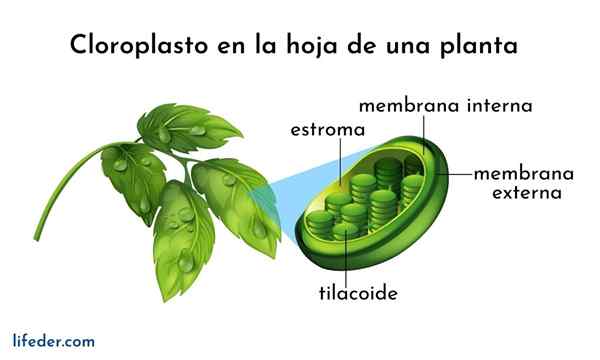
- Tilacoids er et internt tre -dimensjonalt membranholdige kloroplaster. Fullt modne kloroplaster har 40 til 60 grana stablet, med en diameter mellom 0,3 og 0,6 um.
- Antallet tilacoider som utgjør granas varierer mye: fra mindre enn 10 poser i planter utsatt for tilstrekkelig sollys, opp til mer enn 100 tilacoid i planter som lever i miljøer med ekstrem skygge.
- Stablede tilacoider er koblet til hverandre og danner et kontinuerlig rom i kloroplast. Det indre av tilacoid er et ganske romslig rom av vandig natur.
- Tilacoidmembranen er uunnværlig for fotosyntesen, siden den første fasen av prosessen finner sted der.
Tilacoidstruktur
Tilacoider er strukturene som dominerer i en helt moden kloroplast. Hvis en kloroplast vises i det tradisjonelle optiske mikroskopet, kan arter av korn observeres.
Disse er tilacoid stabling; Derfor ble de første observatørene av disse strukturene kalt "Grana".
Ved hjelp av det elektroniske mikroskopet kunne bildet utvides, og det ble konkludert med at arten av disse kornene virkelig var stablede tilacoider.
Dannelsen og strukturen til tilacoidmembranen avhenger av dannelsen av kloroplast fra et ennå ikke differensiert plastidium, kjent som protoplastidio. Tilstedeværelsen av lys stimulerer konverteringen til kloroplaster, og deretter dannelsen av stablede tilacoider.
Kan tjene deg: Sæd: funksjoner, deler, livssyklusTilacoidmembran
I kloroplaster og cyanobakterier er ikke tilacoidmembranen i kontakt med indre plasmamembrandel. Imidlertid begynner Tilacoid -membranformasjonen med invaginering av den indre membranen.
I cyanobakterier og i visse arter av alger blir tilacoider dannet av et enkelt lag med lamella. I kontrast er det et mer komplekst system som finnes i modne kloroplaster.
I denne siste gruppen kan du skille to viktige deler: Grana og Stroma Lamella. Den første består av små stablede album, og den andre er ansvarlig for å koble disse stablingene med hverandre, og danner en struktur fortsetter: Lumen of the Tilacoid.
Lipidsammensetning av membranen
Lipidene som utgjør membranen er høyt spesialiserte og består av nesten 80 % av diacylglycerol galaktosyl: monogalaktosyldiacylglycerol og diglactosyl -diacylglycerol. Disse galaktolipidene har svært umettede kjeder, typiske for tilacoider.
Tilsvarende inneholder tilacoidmembranen lipider som fosfatidylglycerol i mindre proporsjon. Lipidene som er nevnt er ikke fordelt homogent i begge lagene av membranen; Det er en viss grad av asymmetri som ser ut til å bidra til driften av strukturen.
Membranproteinsammensetning
Fotosystemer I og II er de dominerende proteinkomponentene i denne membranen. De er assosiert med cytokrom B -komplekset6F og ATP -syntetase.
Det har blitt funnet at flertallet av elementene i Fotosystem II er lokalisert i de pakket grana -membranene, mens fotosystem I stort sett ligger i de uantilerte tilacoidmembranene. Det vil si at det er en fysisk separasjon mellom begge fotosystemene.
Disse kompleksene inkluderer omfattende membranproteiner, perifere proteiner, kofaktorer og en rekke pigmenter.
Tilacoid lumes
Det indre av tilacoid består av et vandig og tykt stoff, hvis sammensetning er forskjellig fra stroma. Delta i fotofosforylering, lagre protonene som vil generere proton-motorisk kraft for ATP-syntese. I denne prosessen kan pH i lumen nå 4.
I lummen proteom av modellorganismen Talian Arabidopsis Mer enn 80 proteiner er blitt identifisert, men funksjonene deres er ikke fullstendig belyst.
Lumenproteiner er involvert i reguleringen av tilacoid biogenese og i aktivitet og erstatning av proteinene som danner de fotosyntetiske kompleksene, spesielt Photosystem II og NAD (P) H D edrogensa.
Det kan tjene deg: Promielocito: Kjennetegn, funksjoner og patologierTilacoidfunksjoner
Prosessen med fotosyntese, viktig for grønnsaker, begynner i tilacoider. Membranen som avgrenser dem med kloroplaststromen.
Stadier av fotosyntese
Fotosyntese kan deles inn i to store stadier: lysreaksjoner og mørke reaksjoner.
Som navnet tilsier, kan reaksjonene som tilhører den første gruppen bare fortsette i nærvær av lys, mens de i den andre gruppen kan oppstå med eller uten dette. Merk at det ikke er nødvendig at miljøet er "mørkt", det er bare uavhengig av lys.
Den første gruppen av reaksjoner, "lyset", forekommer i tilacoid og kan oppsummeres som følger: Lys + klorofyll + 12 H2O + 12 NADP+ + 18 ADP + 18 PYo à 6 o2 + 12 NADPH + 18 ATP.
Den andre gruppen av reaksjoner forekommer i stroma i kloroplasten og tar ATP og NADPH syntetisert i det første trinnet for å redusere karbondioksidkarbon til glukose (C6H12ENTEN6). Det andre trinnet kan oppsummeres i: 12 NADPH + 18 ATP + 6 CO2 à c6H12ENTEN6 + 12 NADP+ + 18 ADP + 18 PYo + 6 timer2ENTEN.
Lysavhengig stadium
Lysreaksjonene involverer en serie strukturer kjent som fotosystemer, som finnes i tilacoidmembranen og inneholder inne i 300 pigmentmolekyler, blant disse klorofyllene.
Det er to typer fotosystem: den første har en maksimal lysabsorpsjonstopp på 700 nanometer og er kjent som P700, Mens den andre kalles P680. Begge er integrert i tilacoidmembranen.
Prosessen begynner når et av pigmentene absorberer et foton og dette "spretter" mot andre pigmenter. Når et klorofyllmolekyl absorberer lys, absorberer et elektron og et annet molekyl. Molekylet som mistet elektronet er nå oksidert og har en negativ belastning.
P680 Fang lett energi fra klorofyll a. I dette fotosystemet blir et elektron kastet inn i et mer energisystem til en primær elektronakseptor.
Dette elektronet faller til Fotosystem I, gjennom elektrontransportørkjeden. Dette oksidasjons- og reduksjonsreaksjonssystemet er ansvarlig for å overføre protoner og elektroner fra ett molekyl til et annet.
Kan tjene deg: lymfoblasterMed andre ord, det er en strøm av elektroner fra vann til fotosystem II, til fotosystem I og NADPH.
Fotofosforylering
En del av protonene som genereres av dette reaksjonssystemet er lokalisert inne i tilacoid (også kalt Tilacoid Luz), og skaper en kjemisk gradient som genererer en proton-motorisk kraft.
Protonene beveger seg fra tilacoidrommet til stroma, gunstig etter den elektrokjemiske gradienten; det vil si at de forlater tilacoid.
Imidlertid er passering av protoner ikke noe sted i membranen, de må gjøre det gjennom et komplekst enzymatisk system som kalles ATP -synthesase.
Denne protonbevegelsen mot Strom. ATP -syntese ved bruk av lys kalles fotofosforylering.
Disse nevnte stadiene forekommer samtidig: Fotosystem II klorofyll mister et elektron og må erstatte det med et elektron fra brudd på et vannmolekyl; Fotosystemet jeg fanger lys, oksiderer og lanserer et elektron som blir fanget av NADP+.
The Lost Electron of Photo. Disse forbindelsene vil bli brukt i de påfølgende karbonfikseringsreaksjonene, i Calvin -syklusen.
Utvikling
Utviklingen av fotosyntesen som en oksygen -liberende prosess tillot liv slik vi kjenner det.
Det foreslås at fotosyntesen ble utviklet for noen milliarder år siden i stamfaren som ga opphav til nåværende cyanobakterier, basert på et angst fotosyntetisk kompleks.
Det foreslås at utviklingen av fotosyntesen ble ledsaget av to uunnværlige hendelser: opprettelsen av fotosystem p680 og genesen av et indre membransystem, uten tilknytning til cellemembranen.
Det er et essensielt VIPP1 -protein for dannelse av Tilacoides. Dette proteinet er faktisk til stede i planter, alger og cyanobakterier, men fraværende i bakterier som utfører anoksisk fotosyntese.
Det antas at dette genet kan stamme fra gentuplikasjon i den mulige stamfaren til cyanobakterier. Det er bare en cyanobakteriell sak som er i stand til å utføre oksygenfotosyntese og ikke har tilacoides: arten Gloeobacter Violaceus.