

Enolasestruktur, virkningsmekanisme, funksjoner
- 882
- 135
- Thomas Karlsen
De Enolase Det er enzymet som har ansvaret for å utføre konvertering av D-2-fosfoglyserat (2PGA) i fosfoenolpyruvat (PEP) i glykolyse og omvendt reaksjon i glukoneogenese, to metabolske ruter som er en del av den cellulære energimetabolismen.
Beslutningen om å katalysere denne reaksjonen i en eller annen retning avhenger av tilgangen som cellen til glukose har. Det vil si at behovene for å tilpasse stoffskiftet til nedbrytning eller syntese for å oppnå energi. Uunnværlig for realisering av de viktige prosessene.
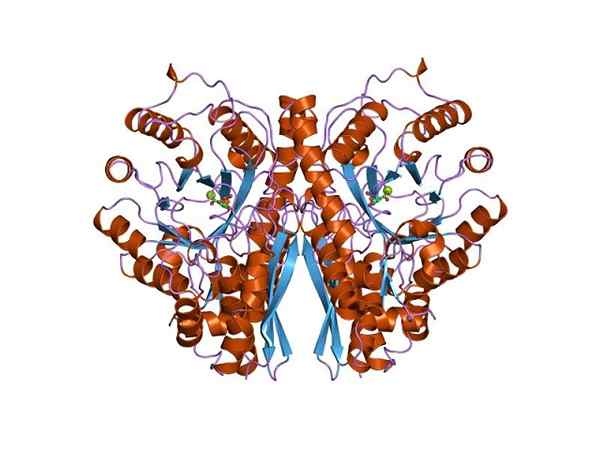
Tre -dimensjonal struktur av enolasa. Av Jawahar Swaminathan og MSD -ansatte ved European Bioinformatics Institute [Public Domain (https: // CreativeCommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons. Siden begge metabolske ruter tilhører sentrum av det sentrale metabolske treet av levende vesener, er det ikke rart at aminosyresekvensen til dette proteinet er bevart i archeae, bakterier og eukaryoter. Og derfor presenterer det lignende katalytiske egenskaper.
Plasseringen av enolase i cellen er begrenset til cytosol, et rom der både glykolyse (også kalt glykolyse) og glukoneogenese i de fleste organismer finner sted.
Imidlertid er det også blitt påvist i andre cellekommer som plasmamembran fra mange kreftceller og celler. Der ser det ut til å være involvert i tilrettelegging av celleformidlingsprosesser, en helt annen funksjon fra den klassiske funksjonen.
Enzymer som er i stand til å utføre mer enn en funksjon, som enolasen gjør, er kjent som måneskinnende enzymer.
[TOC]
Struktur
Linusens kvartærstruktur eller ikke i ligandene er blitt bestemt i et stort antall prokaryotiske og eukaryote individer.
Hver monomer presenterer to domener: et lite amino-terminalt domene og et større karboksylterminal domene. Det N-terminale domenet består av tre α og fire β-ark. Mens C-terminalen består av åtte β-ark som veksler mellom dem som danner en β-tønne som er omgitt av åtte α-propeller.
I tillegg finnes to fagforeningssteder for divalente kationer i hver monomer som har blitt kalt "konformasjonssted" og "katalytisk sted". Den første er ikke veldig selektiv og kan bli med i et bredt utvalg av divalente kationer i fravær av underlag.
Kan tjene deg: sfingolipider: hva er, egenskaper, funksjoner, synteseMens det andre binder seg til ionene etter at underlaget har sammenføyet enzymet. Foreningen av ioner til begge stedene er avgjørende for reaksjonen å fortsette.
Til slutt er det viktig å nevne at i homodimerene blir monomerene sammen med ved å opprettholde en parallell orientering. Derfor er det aktive stedet begrenset til den sentrale regionen dannet av nevnte union.
Imidlertid er det bare en av de to monomerer som deltar i katalyse. Dette forklarer at monomers evne til å utføre reaksjonen under eksperimentelle forhold.
Virkningsmekanismen
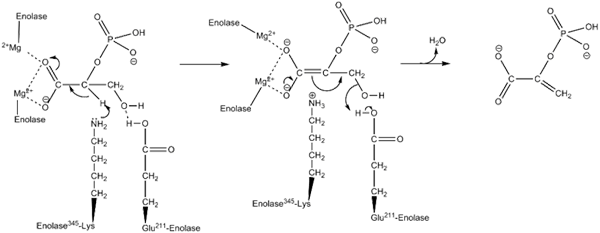 Virkningsmekanisme brukt av enzymet inolase. Av Kthompson08 på engelsk Wikipedia [Public Domain (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons.
Virkningsmekanisme brukt av enzymet inolase. Av Kthompson08 på engelsk Wikipedia [Public Domain (https: // creativecommons.Org/lisenser/by-SA/4.0)], fra Wikimedia Commons. Strukturstudier, så vel som de som har tillatt å bestemme de kinetiske og fysisk -kjemiske egenskapene til enolasa, har tillatt oss å forstå deres virkningsmekanisme.
Måten enzymet utfører katalysen av reaksjonen er ganske interessant. Selv om et enkelt underlag er involvert, er en ordnet sekvensiell mekanisme den som er foreslått.
Dette begynner med foreningen av et ion av Mg2+ til konformasjonsstedet til en av monomerene. Kontinuerlig med foreningen av underlaget til det aktive stedet etterfulgt av forening av et andre ion til det katalytiske stedet og avsluttes med hurtig frigjøring av produktet når reaksjonen er utført. På dette tidspunktet forblir Mg2+ knyttet til konformasjonsstedet.
Langs de samme linjene, for å favorisere realiseringen av reaksjonen det gjennomsnittlige enzymet i første omgang generering av en karbanion -mellomledd, og eliminerer et proton av karbon 2 av 2PGA. Dette gjør det takket være handlingen fra en grunnleggende aminosyrerester.
Sekvensielt foregår karbon 3 hydroksylfjerning ved virkningen av en sur rest fra enzymet. På dette tidspunktet blir foreningen av begge karbonatomer utført ved hjelp av en dobbeltbinding som danner PEP. På denne måten er reaksjonen kulminert.
Det kan tjene deg: Monera Kingdom: Egenskaper, klassifisering og eksemplerFunksjoner
Mange av enzymene så langt studert er i stand til å utføre et bredt utvalg av funksjoner som ikke er relatert til deres "klassiske funksjon" i forskjellige cellulære rom. Disse enzymene har blitt kalt "måneskinn" enzymer.
I denne forstand kan enolase betraktes som et enzym måneskinn, siden mange funksjoner i motsetning til deres klassiske funksjon har blitt tilskrevet til dags dato både i bakterier og eukaryoter.
Noen av disse funksjonene er følgende:
- Delta i vedlikehold av celleform så vel som i vesikulær trafikk når du samhandler med cytoskjelettproteiner.
- I kjernen av pattedyr fungerer som en transkripsjonsfaktor som regulerer uttrykket av gener assosiert med celleproliferasjon. Samarbeide om å opprettholde stabiliteten til RNM i nedbrytningen i bakterier.
- I patogener, for eksempel Streptococcus pneumoniae og Trypanosoma Cruzi, Det ser ut til at det fungerer som en viktig virulensfaktor.
- Det har også blitt funnet det i Streptococcus pyogenes, Enolase skilles ut til det ekstracellulære medium som letter vevsnedbrytning og unnvikelsen av vertssystemet.
- På overflaten av tumorceller uttrykkes det ved å styrke metastasen.
Enolase og dets forhold til celledidsmekanismer
Tallrike patogener, så vel som tumorceller, uttrykker i membranen eller skiller ut de ekstracellulære medium proteaser som er i stand til å nedbryte proteinproteiner.
Denne kapasiteten lar disse cellene bryte gjennom vevene og spre seg raskt gjennom hele vertsorganismen. Dermed favoriserer unndragelsen av immunforsvaret og derfor etablering av infeksjonen.
Selv når enolasa mangler proteaseaktivitet, deltar du i prosessen med å spre mange patogener i verten så vel som tumorceller under målstastase.
Dette oppnår det takket være det faktum at det kommer til uttrykk på overflaten av disse cellene som fungerer som en plasminogenreseptor. Det siste er zimogenet til en serin-protoTease kjent som plasminat som er en del av det fibrinolytiske systemet og virker nedbrytende ekstracellulære matriksproteiner.
Derfor er enolasen uttrykt på overflaten en strategi som disse cellene har skaffet seg for å etablere infeksjonen og spredte vellykket.
Det kan tjene deg: Sympatrisk spesifikasjon: Konsept, egenskaper og eksemplerDenne strategien består av to prosesser:
- Unnvikelsen av vertens immunforsvar. Siden disse cellene fra et vertsprotein er belagt, blir de ignorert av immunsystemceller som gjenkjenner ikke -ned -patogener assosiert med patogener.
- Forlengning av plasminogen etter handlinger i plasminat. Hvis deltakelse i nedbrytningen av ekstracellulære matriksproteiner, letter deretter den raske og effektive formidlingen.
Referanser
- Avilan L, Gualdron-Lopez M, Quiñones W, González-González L, Hannaert V, Michels PAA, Concepción JL. Enolase: En nøkkelspiller i metabolismen og en sannsynlig virulensfaktor av trypanosomatidparasitter-PerSpectives for bruk som et terapeutisk mål. Enzymforskning. 2011 Vol. Artikkel ID932549, 14 sider.
- Bhowmick I, Kumar N, Sharma S, Coppens I, Jarori GK, Plasmodium falciparum enosoase: scenespesifikk uttrykk og subcellulær lokalisering. Malaria Journal. 2009; 8 (1). Artikkel 179.
- Dag I, Peshavaria M, Quinn GB, en differensiell molekylær klokke i enoase isoprotein evolusjon. Journal of Molecular Evolution. 1993; 36 (6): 599-601.
- av Torre-Scuder E, Manzano-Román R, Pérez-Sánchez R, Schiles-Lucas M, Oleaga A. Kloning og karakterisering av plasminogenbindende overflate -ssosiert enolaase fra Schistosoma Bovis. Veterinærparasitologi. 2010; 173: 73-84.
- Dinovo EC, Boyer PD. Isotopiske sonder av enolase -reaksjonsmekanismen. Opprinnelige og equiquibium -isotoputvekslingskurser: Primær- og Secandary Isotope -effekter. J Biol Chem. 1971; 246 (14): 4586-4593.
- Kaberdin VR, Lin-Chao S, Å avdekke nye roller for mindre komponenter i E. Coli RNA nedbrytning. RNA -biologi. 2009; 6 (4): 402-405.
- Keller A, Peltzer J, Carpentier G. Interaksjoner mellom enolas. Biochimica et Biophysica Acta. 2007; 1770 (6): 919-926.
- Lung J, Liu KJ, Chang JY, Leu SJ, Shih NY. MBP-1 er effektivt kodet av et alternativt transkripsjon av ENRI1-genet Buttranslationalt regulert av proteasomavhengig proteinforskyvning. FEBS Journal. 2010; 277 (20): 4308-4321.
- Pancholi v. Multifunksjonell a-enolase: dens rolle i sykdommer. Cellular og Molecular Life Sciences. 2001; 58 (7): 902-920.
- Poyner RR, Cleland WW, Reed GH. Rollen til metallinnlegg i katalyse ved enolase. En ordnet kinetisk mekanisme for et enkelt underlag enzym. Biokjemi. 2001; 40: 9008-8017.
- Segovia-Gamboa NC, Chávez-Munguía B, Medina-Flores A, Entamoeba invadens, Encystation prosess og enolase. Eksperimentell parasitologi. 2010; 125 (2): 63-69.
- Tanaka M, Sugisaki K, Nakashima K, bytter i nivåer av oversettbare mRNA for enoase isozymer under utvikling av kyllingskjelettmuskel. Biokjemisk og biofysisk forskningskommunikasjon. 1985; 133 (3): 868-872.